

Songbird: a unique animal model for studying the molecular basis of disorders of vocal development and communication
- PMID: 25912323
- PMCID: PMC4547995
- DOI: 10.1538/expanim.15-0008
Songbird: a unique animal model for studying the molecular basis of disorders of vocal development and communication
Abstract
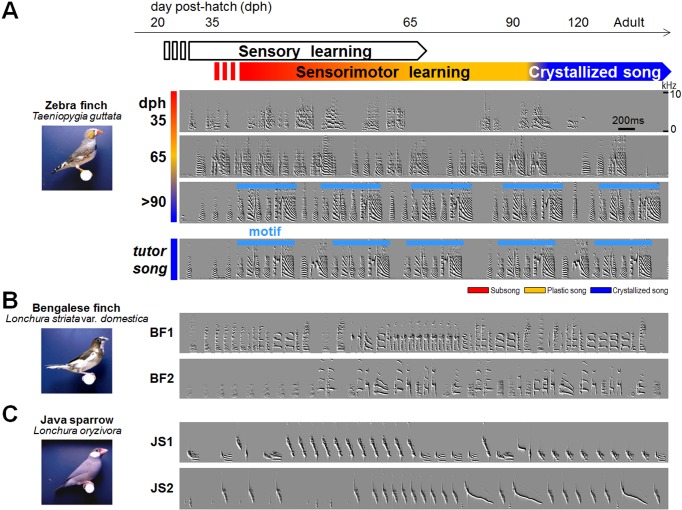
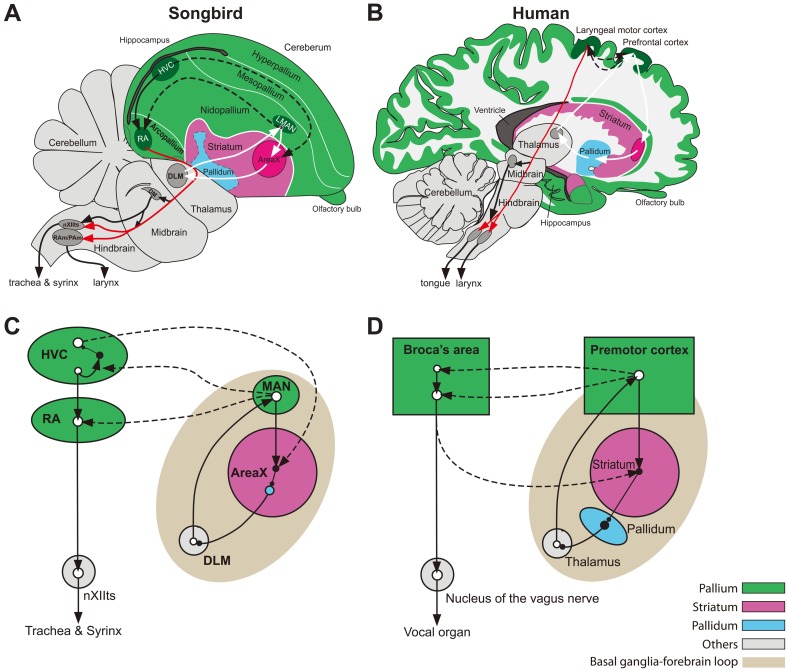
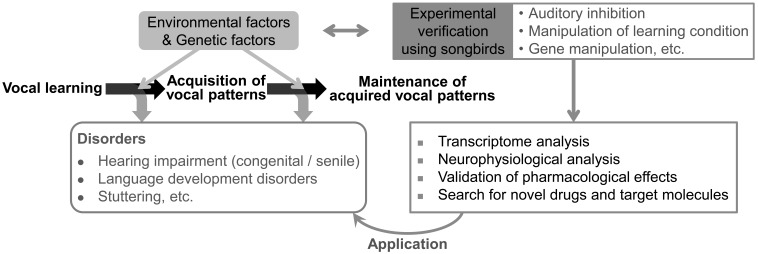
Like humans, songbirds are one of the few animal groups that learn vocalization. Vocal learning requires coordination of auditory input and vocal output using auditory feedback to guide one's own vocalizations during a specific developmental stage known as the critical period. Songbirds are good animal models for understand the neural basis of vocal learning, a complex form of imitation, because they have many parallels to humans with regard to the features of vocal behavior and neural circuits dedicated to vocal learning. In this review, we will summarize the behavioral, neural, and genetic traits of birdsong. We will also discuss how studies of birdsong can help us understand how the development of neural circuits for vocal learning and production is driven by sensory input (auditory information) and motor output (vocalization).
Figures

Similar articles
-
Developmental regulation of basal ganglia circuitry during the sensitive period for vocal learning in songbirds.Ann N Y Acad Sci. 2004 Jun;1016:395-415. doi: 10.1196/annals.1298.037. Ann N Y Acad Sci. 2004. PMID: 15313787 Review.
-
Shared mechanisms of auditory and non-auditory vocal learning in the songbird brain.Elife. 2022 Sep 15;11:e75691. doi: 10.7554/eLife.75691. Elife. 2022. PMID: 36107757 Free PMC article.
-
Neural mechanisms for learned birdsong.Learn Mem. 2009 Oct 22;16(11):655-69. doi: 10.1101/lm.1065209. Print 2009 Nov. Learn Mem. 2009. PMID: 19850665 Review.
-
Variability in action: Contributions of a songbird cortical-basal ganglia circuit to vocal motor learning and control.Neuroscience. 2015 Jun 18;296:39-47. doi: 10.1016/j.neuroscience.2014.10.010. Epub 2014 Oct 18. Neuroscience. 2015. PMID: 25445191 Review.
-
Auditory feedback in learning and maintenance of vocal behaviour.Nat Rev Neurosci. 2000 Oct;1(1):31-40. doi: 10.1038/35036205. Nat Rev Neurosci. 2000. PMID: 11252766 Review.
Cited by
-
Therapeutic Potential of α-Synuclein Evolvability for Autosomal Recessive Parkinson's Disease.Parkinsons Dis. 2021 Nov 23;2021:6318067. doi: 10.1155/2021/6318067. eCollection 2021. Parkinsons Dis. 2021. PMID: 34858569 Free PMC article. Review.
-
Is the Capacity for Vocal Learning in Vertebrates Rooted in Fish Schooling Behavior?Evol Biol. 2018;45(4):359-373. doi: 10.1007/s11692-018-9457-8. Epub 2018 Jun 13. Evol Biol. 2018. PMID: 30459479 Free PMC article. Review.
-
Grand and Less Grand Challenges in Avian Physiology.Front Physiol. 2017 Apr 19;8:222. doi: 10.3389/fphys.2017.00222. eCollection 2017. Front Physiol. 2017. PMID: 28469579 Free PMC article. No abstract available.
-
Does Zipf's law of abbreviation shape birdsong?PLoS Comput Biol. 2025 Aug 13;21(8):e1013228. doi: 10.1371/journal.pcbi.1013228. eCollection 2025 Aug. PLoS Comput Biol. 2025. PMID: 40802653 Free PMC article.
-
Vocal changes in a zebra finch model of Parkinson's disease characterized by alpha-synuclein overexpression in the song-dedicated anterior forebrain pathway.PLoS One. 2022 May 4;17(5):e0265604. doi: 10.1371/journal.pone.0265604. eCollection 2022. PLoS One. 2022. PMID: 35507553 Free PMC article.
References
-
- Ackermann H., Hage S.R., Ziegler W.2014. Brain mechanisms of acoustic communication in humans and nonhuman primates: an evolutionary perspective. Behav. Brain Sci. 37: 529–546. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources