

TEAD and YAP regulate the enhancer network of human embryonic pancreatic progenitors
- PMID: 25915126
- PMCID: PMC4434585
- DOI: 10.1038/ncb3160
TEAD and YAP regulate the enhancer network of human embryonic pancreatic progenitors
Abstract
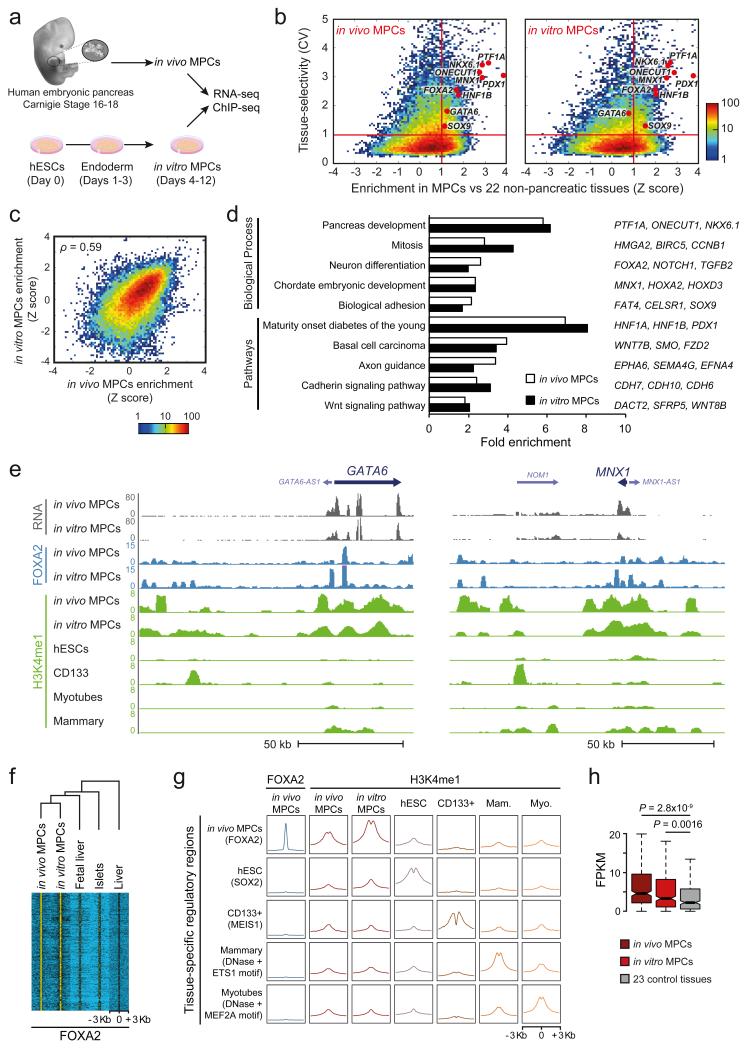
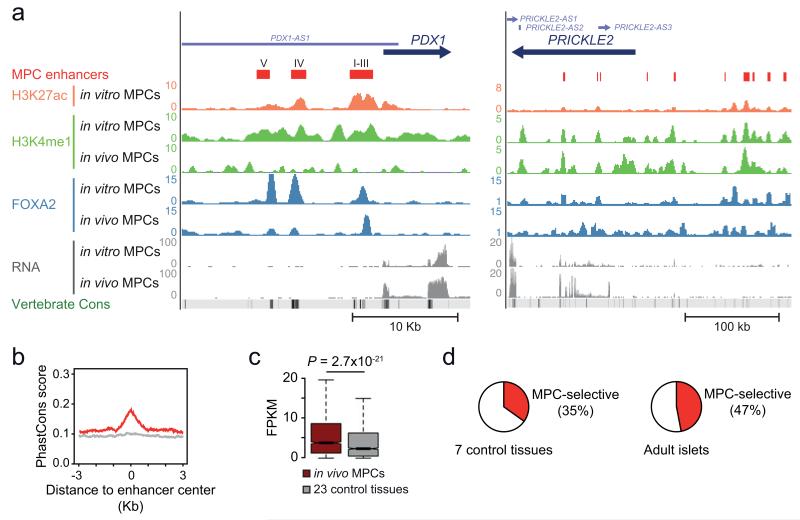
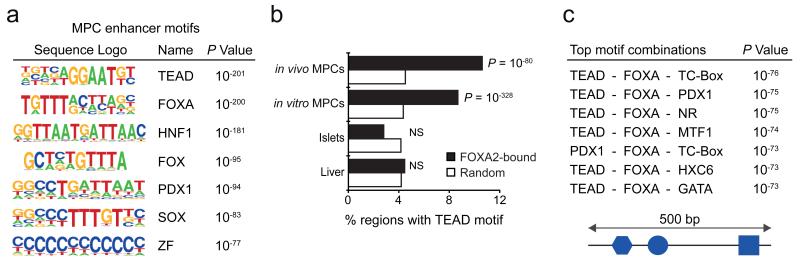
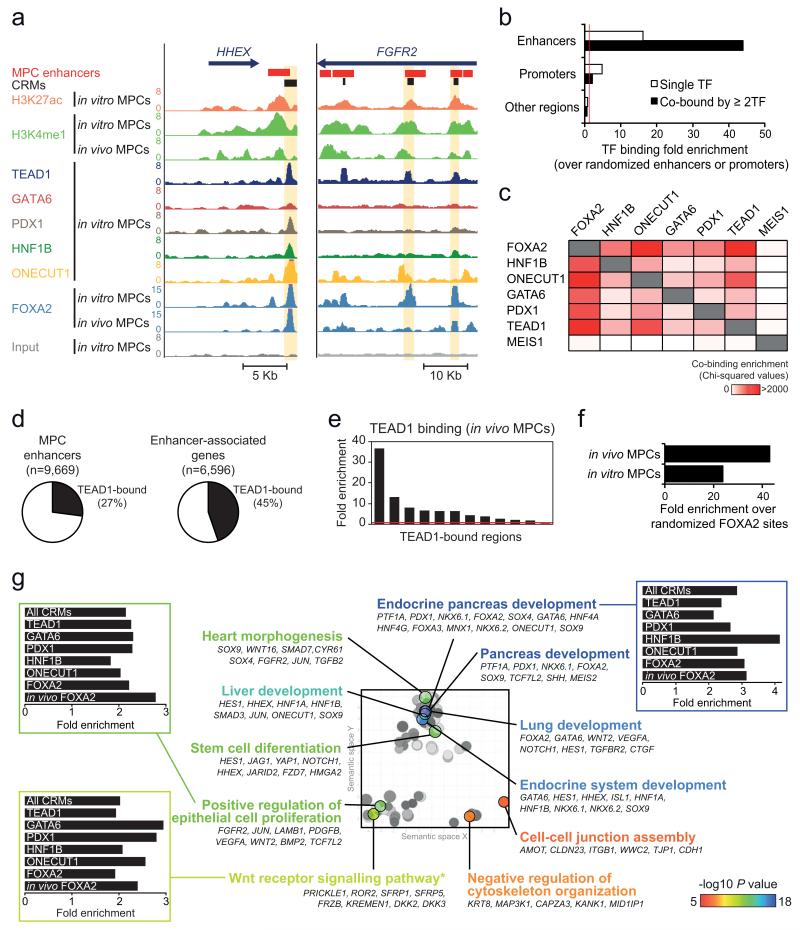
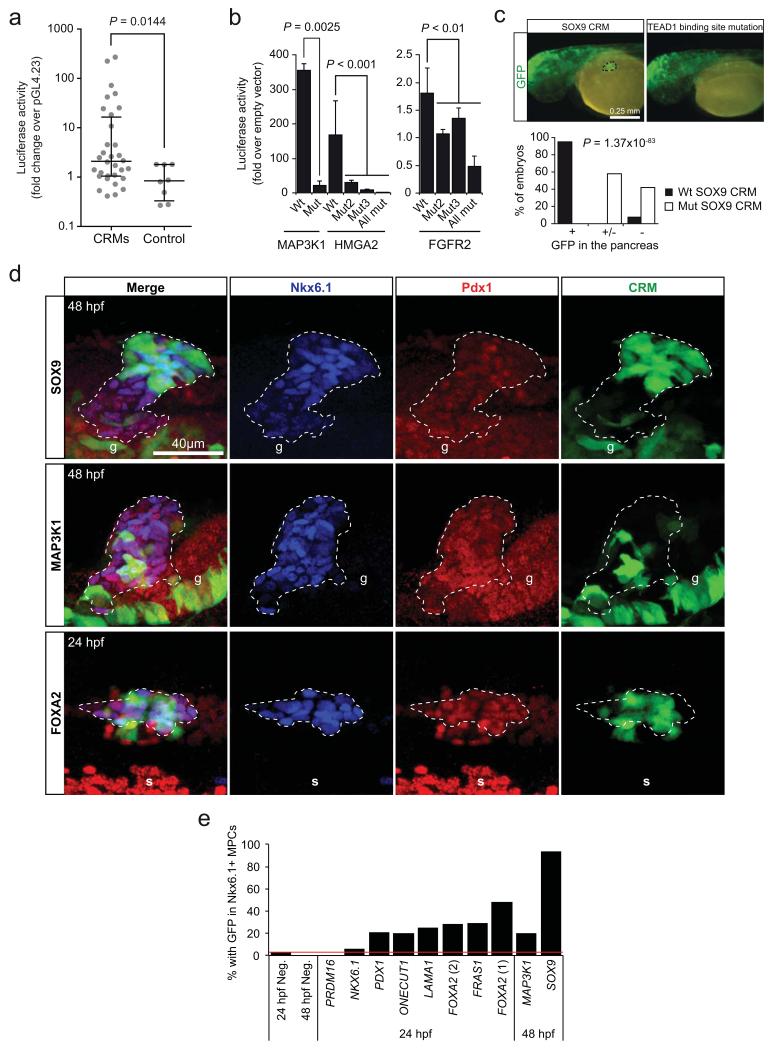
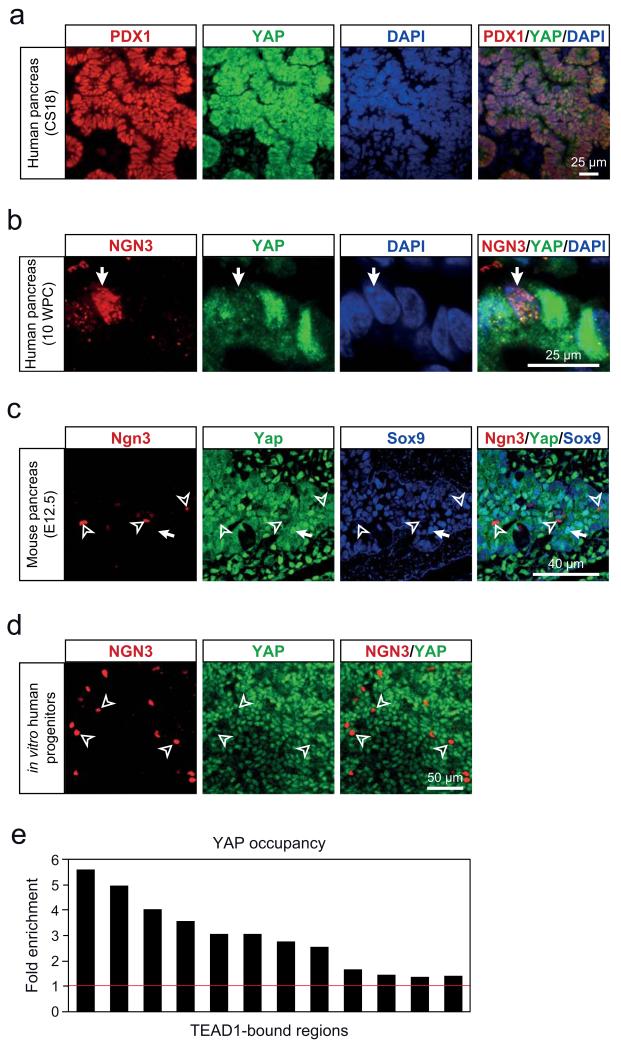
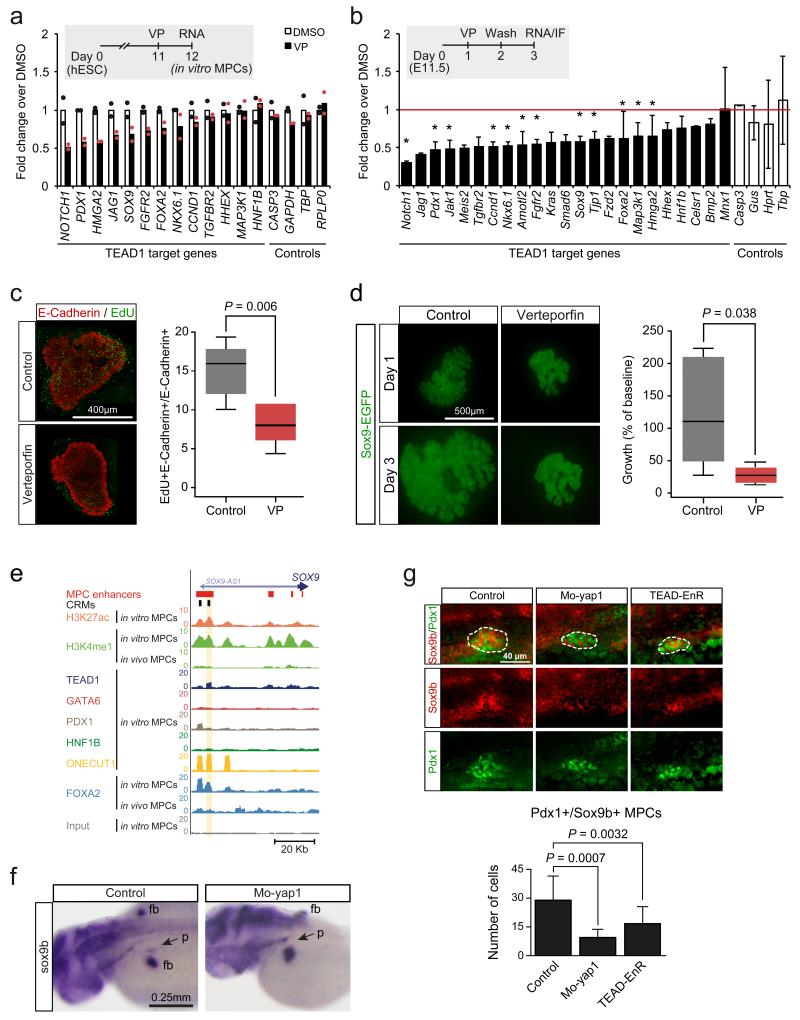
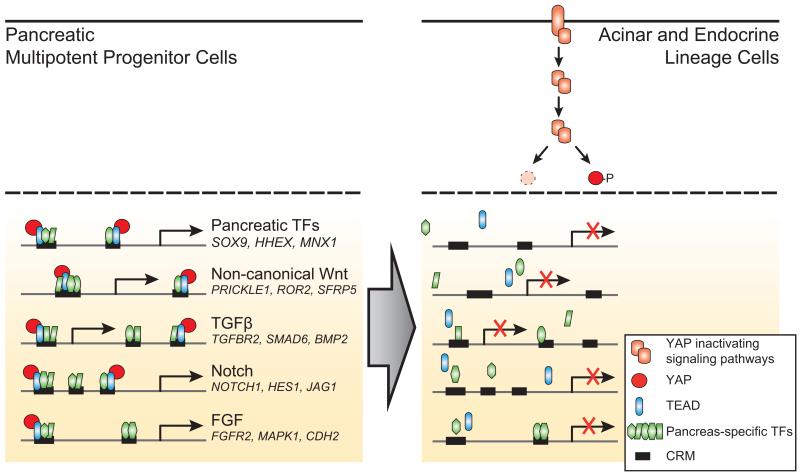
The genomic regulatory programmes that underlie human organogenesis are poorly understood. Pancreas development, in particular, has pivotal implications for pancreatic regeneration, cancer and diabetes. We have now characterized the regulatory landscape of embryonic multipotent progenitor cells that give rise to all pancreatic epithelial lineages. Using human embryonic pancreas and embryonic-stem-cell-derived progenitors we identify stage-specific transcripts and associated enhancers, many of which are co-occupied by transcription factors that are essential for pancreas development. We further show that TEAD1, a Hippo signalling effector, is an integral component of the transcription factor combinatorial code of pancreatic progenitor enhancers. TEAD and its coactivator YAP activate key pancreatic signalling mediators and transcription factors, and regulate the expansion of pancreatic progenitors. This work therefore uncovers a central role for TEAD and YAP as signal-responsive regulators of multipotent pancreatic progenitors, and provides a resource for the study of embryonic development of the human pancreas.
Figures
References
METHODS REFERENCES
-
- O’Rahilly R, Müller F, Hutchins GM, Moore GW. Computer ranking of the sequence of appearance of 73 features of the brain and related structures in staged human embryos during the sixth week of development. Am. J. Anat. 1987;180:69–86. - PubMed
-
- Maestro MA, et al. Hnf6 and Tcf2 (MODY5) are linked in a gene network operating in a precursor cell domain of the embryonic pancreas. Human Molecular Genetics. 2003;12:3307–3314. - PubMed
-
- Piper K, et al. Beta cell differentiation during early human pancreas development. J. Endocrinol. 2004;181:11–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
