

Collecting lymphatic vessel permeability facilitates adipose tissue inflammation and distribution of antigen to lymph node-homing adipose tissue dendritic cells
- PMID: 25917096
- PMCID: PMC4433841
- DOI: 10.4049/jimmunol.1500221
Collecting lymphatic vessel permeability facilitates adipose tissue inflammation and distribution of antigen to lymph node-homing adipose tissue dendritic cells
Abstract
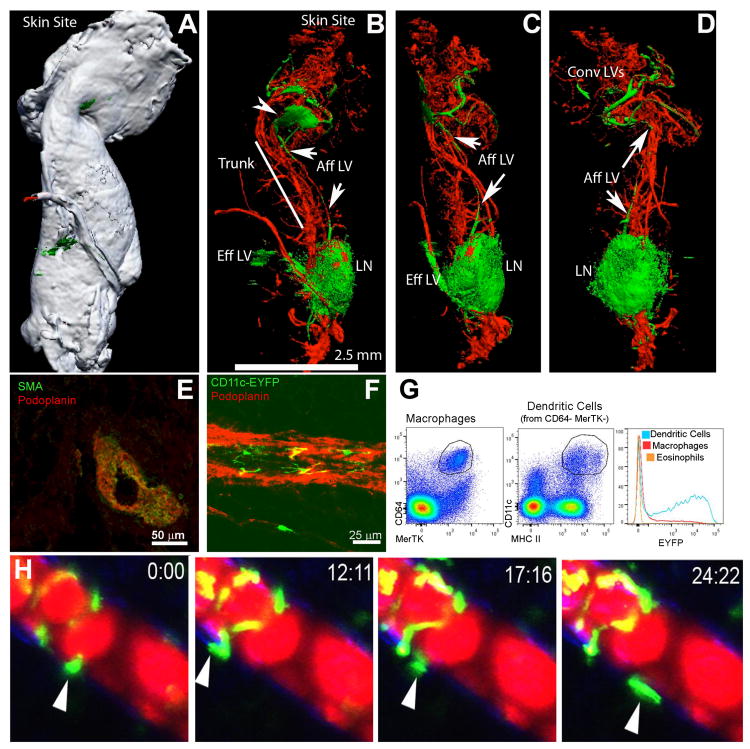
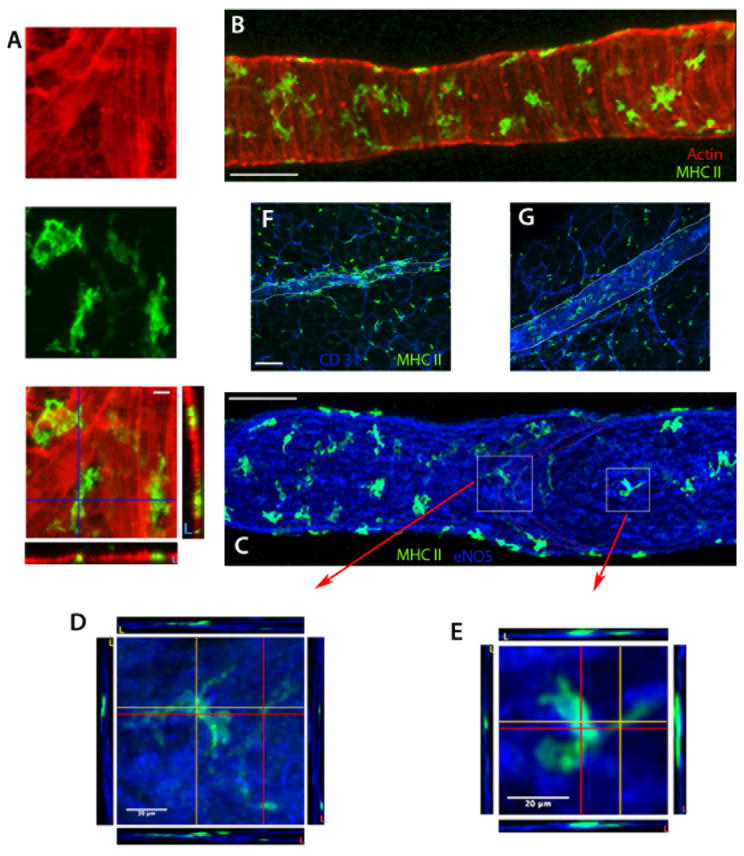
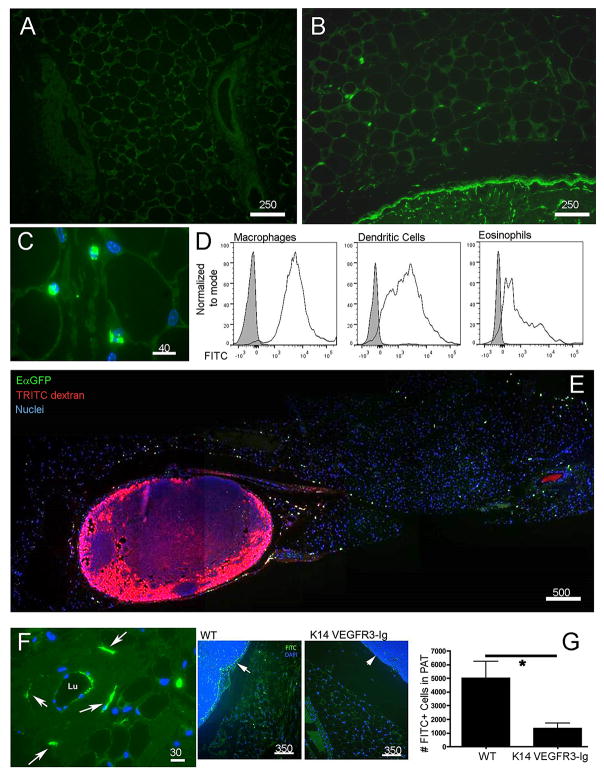
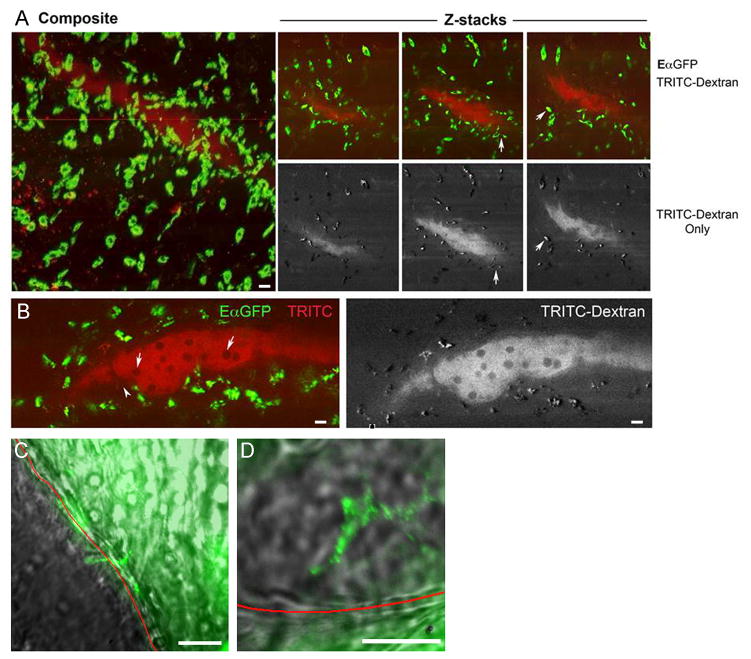
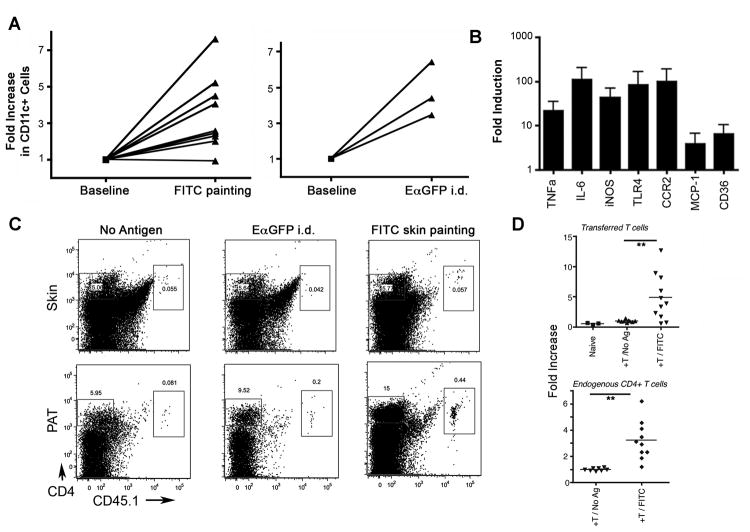
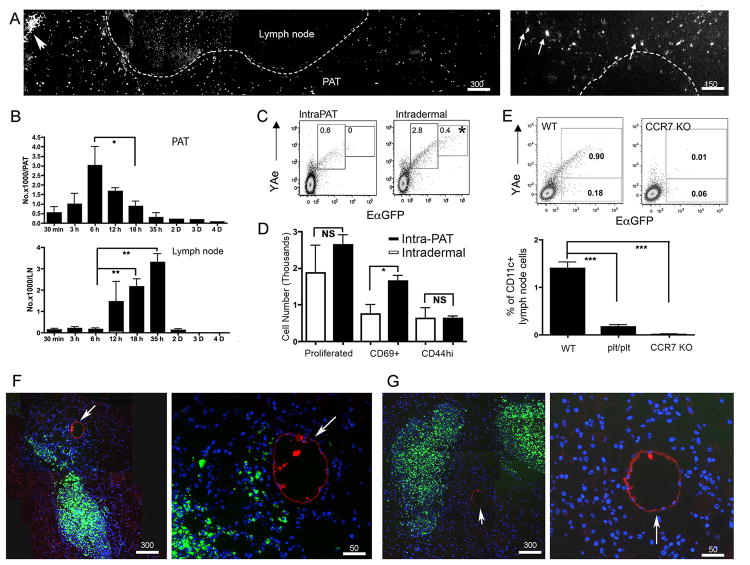
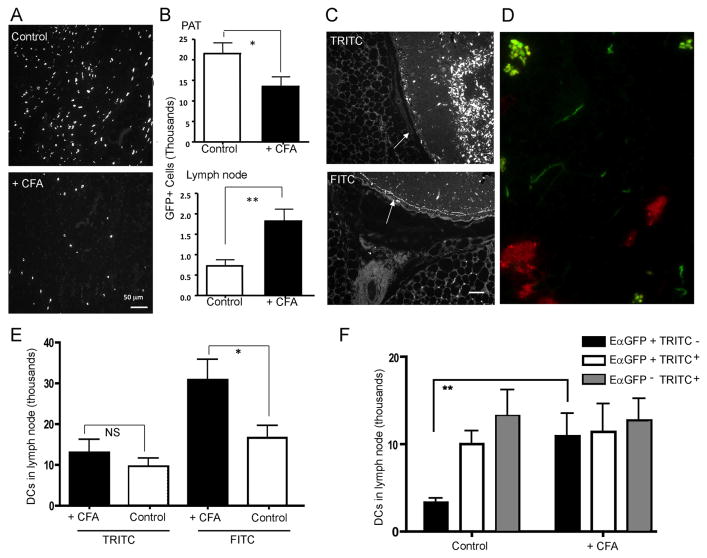
Collecting lymphatic vessels (CLVs), surrounded by fat and endowed with contractile muscle and valves, transport lymph from tissues after it is absorbed into lymphatic capillaries. CLVs are not known to participate in immune responses. In this study, we observed that the inherent permeability of CLVs allowed broad distribution of lymph components within surrounding fat for uptake by adjacent macrophages and dendritic cells (DCs) that actively interacted with CLVs. Endocytosis of lymph-derived Ags by these cells supported recall T cell responses in the fat and also generated Ag-bearing DCs for emigration into adjacent lymph nodes (LNs). Enhanced recruitment of DCs to inflammation-reactive LNs significantly relied on adipose tissue DCs to maintain sufficient numbers of Ag-bearing DCs as the LN expanded. Thus, CLVs coordinate inflammation and immunity within adipose depots and foster the generation of an unexpected pool of APCs for Ag transport into the adjacent LN.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures
References
-
- Schmid-Schonbein GW. Microlymphatics and lymph flow. Physiol Rev. 1990;70:987–1028. - PubMed
-
- Muthuchamy M, Zawieja D. Molecular regulation of lymphatic contractility. Ann N Y Acad Sci. 2008;1131:89–99. - PubMed
-
- Randolph GJ, Angeli V, Swartz MA. Dendritic-cell trafficking to lymph nodes through lymphatic vessels. Nature reviews Immunology. 2005;5:617–628. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI082982/AI/NIAID NIH HHS/United States
- R37 AI049653/AI/NIAID NIH HHS/United States
- HL70308/HL/NHLBI NIH HHS/United States
- HL075199/HL/NHLBI NIH HHS/United States
- R21 AG046743/AG/NIA NIH HHS/United States
- P30 DK05341/DK/NIDDK NIH HHS/United States
- R01 HL081151/HL/NHLBI NIH HHS/United States
- R01 AI055037/AI/NIAID NIH HHS/United States
- R01 AI049653/AI/NIAID NIH HHS/United States
- R01 HL075199/HL/NHLBI NIH HHS/United States
- R21 HL085659/HL/NHLBI NIH HHS/United States
- 100262/Wellcome Trust/United Kingdom
- AI055037/AI/NIAID NIH HHS/United States
- R01 AG030578/AG/NIA NIH HHS/United States
- R01 HL070308/HL/NHLBI NIH HHS/United States
- U01 AI082982/AI/NIAID NIH HHS/United States
- HL081151/HL/NHLBI NIH HHS/United States
- R01 HL084312/HL/NHLBI NIH HHS/United States
- R21 AI055037/AI/NIAID NIH HHS/United States
- R01 HL096552/HL/NHLBI NIH HHS/United States
- HL084312/HL/NHLBI NIH HHS/United States
- HL085659/HL/NHLBI NIH HHS/United States
- AI 049653/AI/NIAID NIH HHS/United States
- AG030578/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
