

Review series: The cell biology of renal filtration
- PMID: 25918223
- PMCID: PMC4411276
- DOI: 10.1083/jcb.201410017
Review series: The cell biology of renal filtration
Abstract
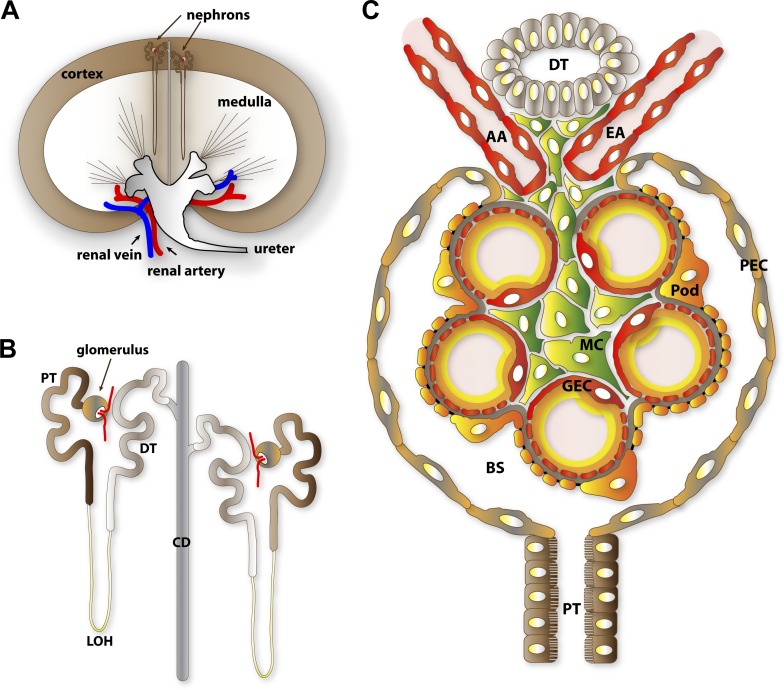
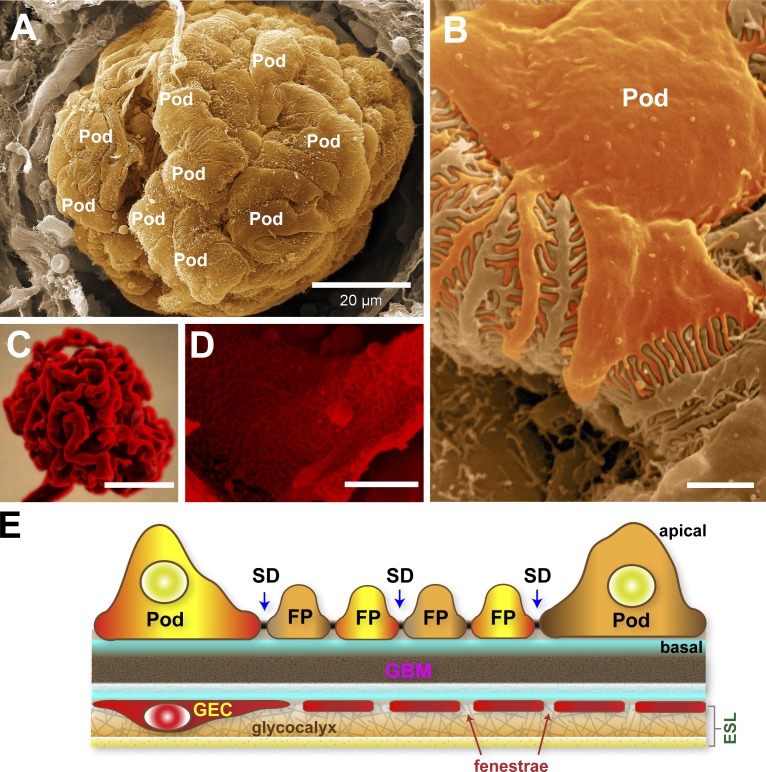
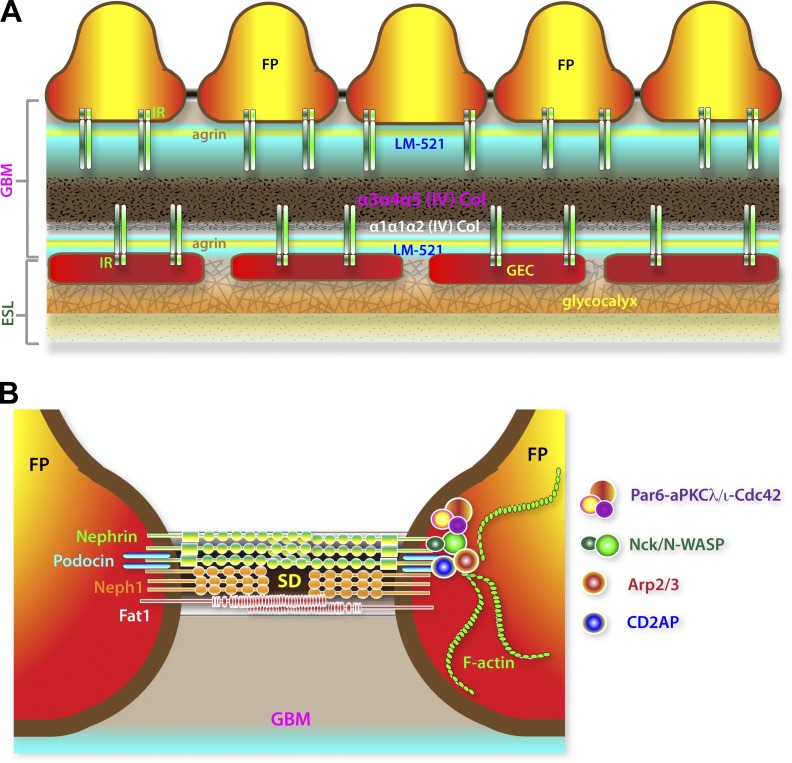
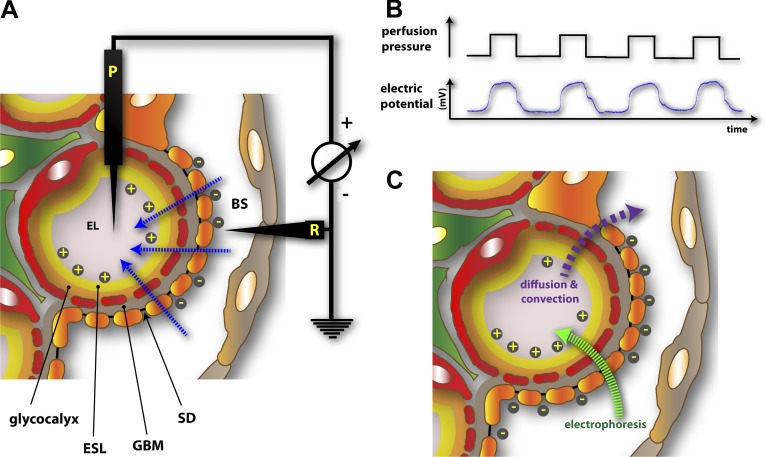
The function of the kidney, filtering blood and concentrating metabolic waste into urine, takes place in an intricate and functionally elegant structure called the renal glomerulus. Normal glomerular function retains circulating cells and valuable macromolecular components of plasma in blood, resulting in urine with just trace amounts of proteins. Endothelial cells of glomerular capillaries, the podocytes wrapped around them, and the fused extracellular matrix these cells form altogether comprise the glomerular filtration barrier, a dynamic and highly selective filter that sieves on the basis of molecular size and electrical charge. Current understanding of the structural organization and the cellular and molecular basis of renal filtration draws from studies of human glomerular diseases and animal models of glomerular dysfunction.
© 2015 Scott and Quaggin.
Figures
References
-
- Akilesh S., Suleiman H., Yu H., Stander M.C., Lavin P., Gbadegesin R., Antignac C., Pollak M., Kopp J.B., Winn M.P., and Shaw A.S.. 2011. Arhgap24 inactivates Rac1 in mouse podocytes, and a mutant form is associated with familial focal segmental glomerulosclerosis. J. Clin. Invest. 121:4127–4137 10.1172/JCI46458 - DOI - PMC - PubMed
-
- Ashraf S., Gee H.Y., Woerner S., Xie L.X., Vega-Warner V., Lovric S., Fang H., Song X., Cattran D.C., Avila-Casado C., et al. 2013. ADCK4 mutations promote steroid-resistant nephrotic syndrome through CoQ10 biosynthesis disruption. J. Clin. Invest. 123:5179–5189 10.1172/JCI69000 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
