

Impact of acute malaria on pre-existing antibodies to viral and vaccine antigens in mice and humans
- PMID: 25919588
- PMCID: PMC4412709
- DOI: 10.1371/journal.pone.0125090
Impact of acute malaria on pre-existing antibodies to viral and vaccine antigens in mice and humans
Abstract
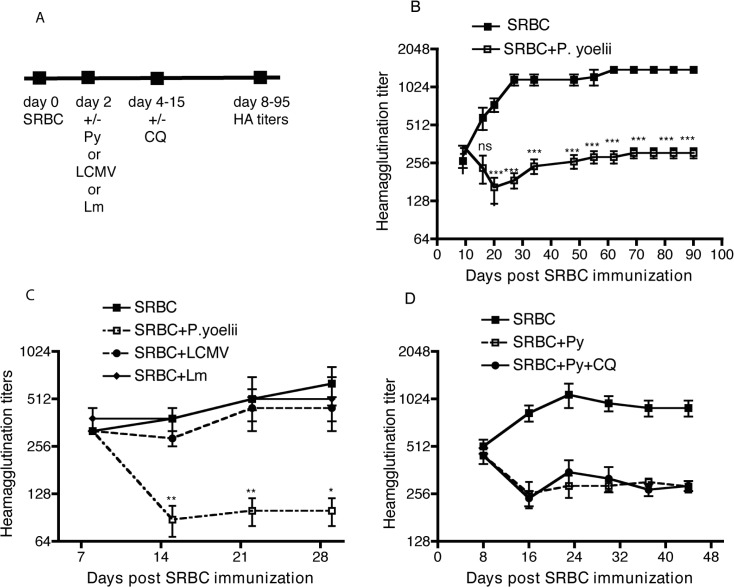
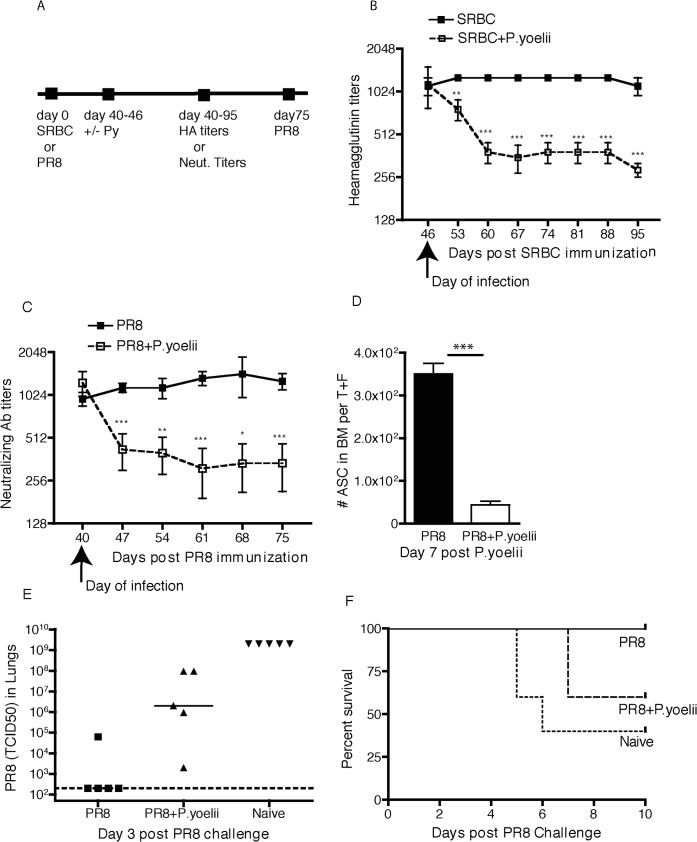
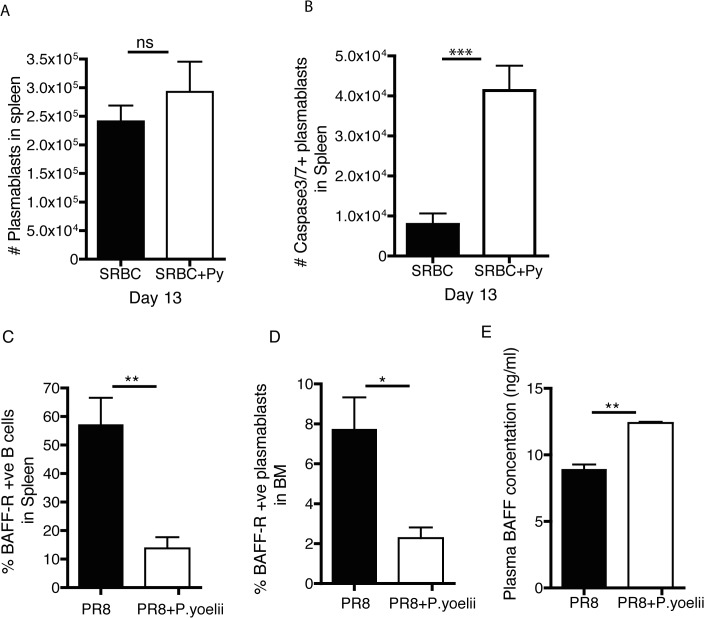
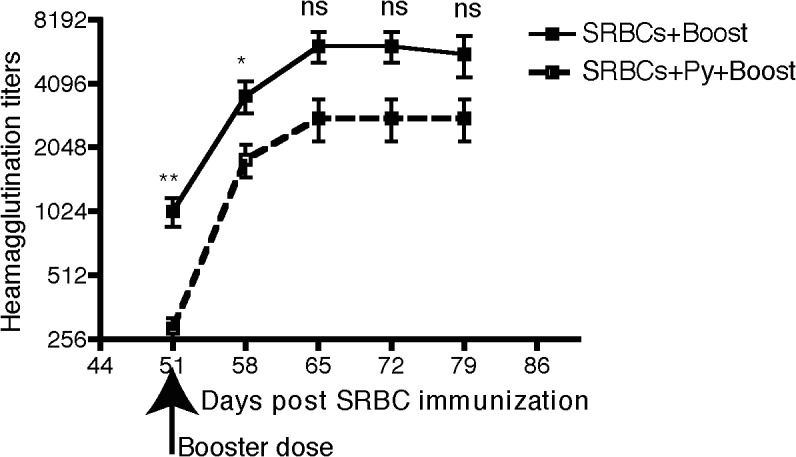
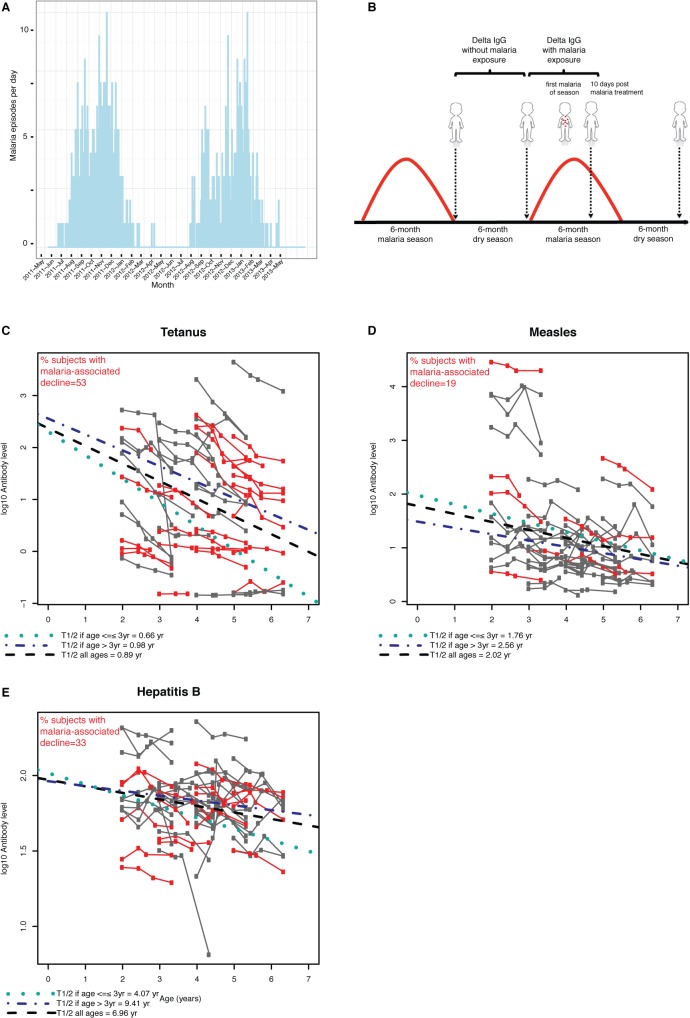
Vaccine-induced immunity depends on long-lived plasma cells (LLPCs) that maintain antibody levels. A recent mouse study showed that Plasmodium chaubaudi infection reduced pre-existing influenza-specific antibodies--raising concerns that malaria may compromise pre-existing vaccine responses. We extended these findings to P. yoelii infection, observing decreases in antibodies to model antigens in inbred mice and to influenza in outbred mice, associated with LLPC depletion and increased susceptibility to influenza rechallenge. We investigated the implications of these findings in Malian children by measuring vaccine-specific IgG (tetanus, measles, hepatitis B) before and after the malaria-free 6-month dry season, 10 days after the first malaria episode of the malaria season, and after the subsequent dry season. On average, vaccine-specific IgG did not decrease following acute malaria. However, in some children malaria was associated with an accelerated decline in vaccine-specific IgG, underscoring the need to further investigate the impact of malaria on pre-existing vaccine-specific antibodies.
Conflict of interest statement
Figures
References
-
- Keja K, Chan C, Hayden G, Henderson RH. Expanded programme on immunization. World health statistics quarterly Rapport trimestriel de statistiques sanitaires mondiales. 1988;41(2):59–63. PubMed . - PubMed
-
- Centers for Disease C, Prevention. Global routine vaccination coverage, 2011. MMWR Morbidity and mortality weekly report. 2012;61(43):883–5. PubMed . - PubMed
-
- Greenwood BM, Bradley-Moore AM, Bryceson AD, Palit A. Immunosuppression in children with malaria. Lancet. 1972;1(7743):169–72. PubMed . - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
