

The control of reactive oxygen species production by SHP-1 in oligodendrocytes
- PMID: 25919645
- PMCID: PMC4534322
- DOI: 10.1002/glia.22842
The control of reactive oxygen species production by SHP-1 in oligodendrocytes
Abstract
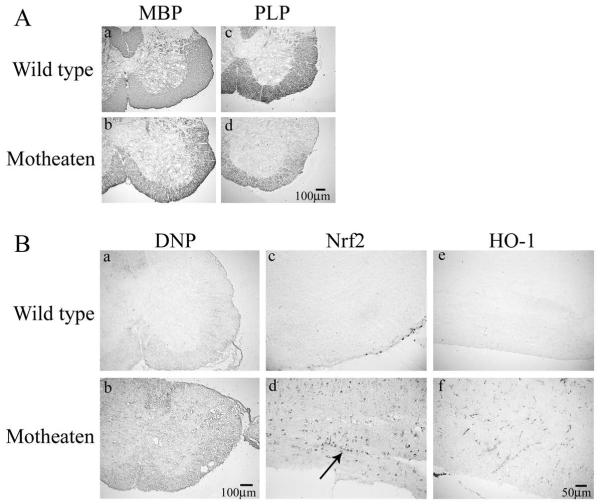
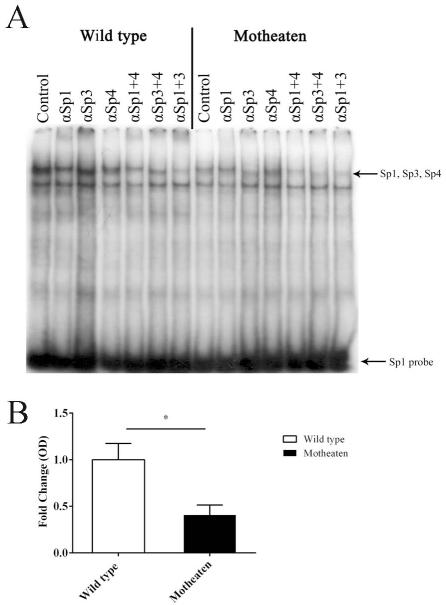
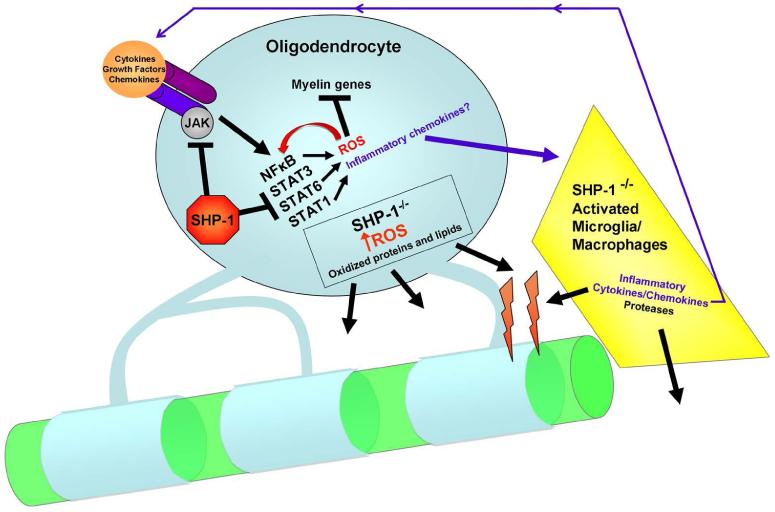
We have previously described reduced myelination and corresponding myelin basic protein (MBP) expression in the central nervous system of Src homology 2 domain-containing protein tyrosine phosphatase 1 (SHP-1) deficient motheaten (me/me) mice compared with normal littermate controls. Deficiency in myelin and MBP expression in both brains and spinal cords of motheaten mice correlated with reduced MBP mRNA expression levels in vivo and in purified oligodendrocytes in vitro. Therefore, SHP-1 activity seems to be a critical regulator of oligodendrocyte gene expression and function. Consistent with this role, this study demonstrates that oligodendrocytes of motheaten mice and SHP-1-depleted N20.1 cells produce higher levels of reactive oxygen species (ROS) and exhibit corresponding markers of increased oxidative stress. In agreement with these findings, we demonstrate that increased production of ROS coincides with ROS-induced signaling pathways known to affect myelin gene expression in oligodendrocytes. Antioxidant treatment of SHP-1-deficient oligodendrocytes reversed the pathological changes in these cells, with increased myelin protein gene expression and decreased expression of nuclear factor (erythroid-2)-related factor 2 (Nrf2) responsive gene, heme oxygenase-1 (HO-1). Furthermore, we demonstrate that SHP-1 is expressed in human white matter oligodendrocytes, and there is a subset of multiple sclerosis subjects that demonstrate a deficiency of SHP-1 in normal-appearing white matter. These studies reveal critical pathways controlled by SHP-1 in oligodendrocytes that relate to susceptibility of SHP-1-deficient mice to both developmental defects in myelination and to inflammatory demyelinating diseases.
Keywords: inflammation; multiple sclerosis; myelin; normal-appearing white matter; oxidative stress.
© 2015 Wiley Periodicals, Inc.
Figures






References
-
- Anrather J, Racchumi G, Iadecola C. NF-kappaB regulates phagocytic NADPH oxidase by inducing the expression of gp91phox. J Biol Chem. 2006;281:5657–67. - PubMed
-
- Berlett BS, Stadtman ER. Protein oxidation in aging, disease, and oxidative stress. J Biol Chem. 1997;272:20313–6. - PubMed
-
- Cammer W. Protection of cultured oligodendrocytes against tumor necrosis factor-alpha by the antioxidants coenzyme Q(10) and N-acetyl cysteine. Brain Res Bull. 2002;58:587–92. - PubMed
-
- Cammer W, Zhang H. Maturation of oligodendrocytes is more sensitive to TNF alpha than is survival of precursors and immature oligodendrocytes. J Neuroimmunol. 1999;97:37–42. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
