

SIRT1-Mediated eNAMPT Secretion from Adipose Tissue Regulates Hypothalamic NAD+ and Function in Mice
- PMID: 25921090
- PMCID: PMC4426056
- DOI: 10.1016/j.cmet.2015.04.002
SIRT1-Mediated eNAMPT Secretion from Adipose Tissue Regulates Hypothalamic NAD+ and Function in Mice
Abstract
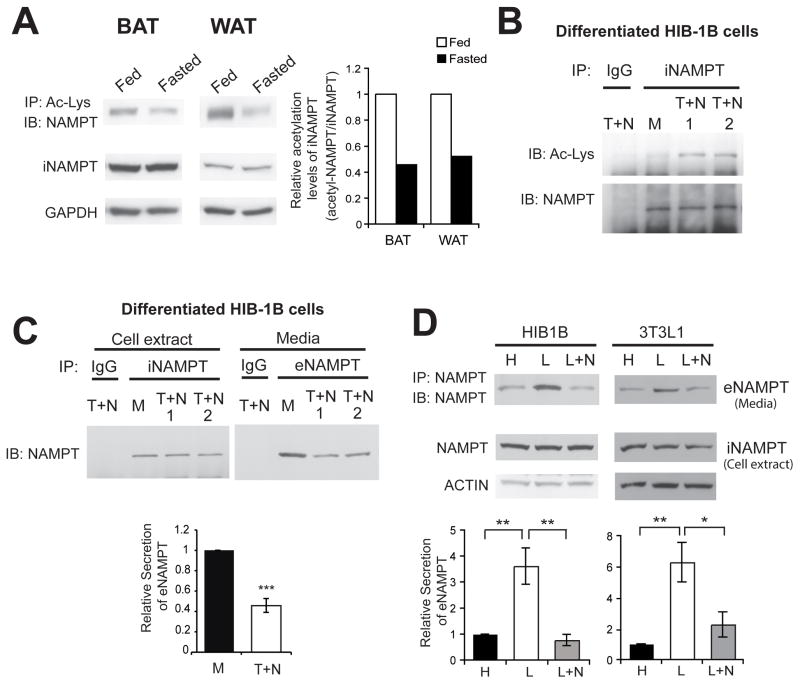
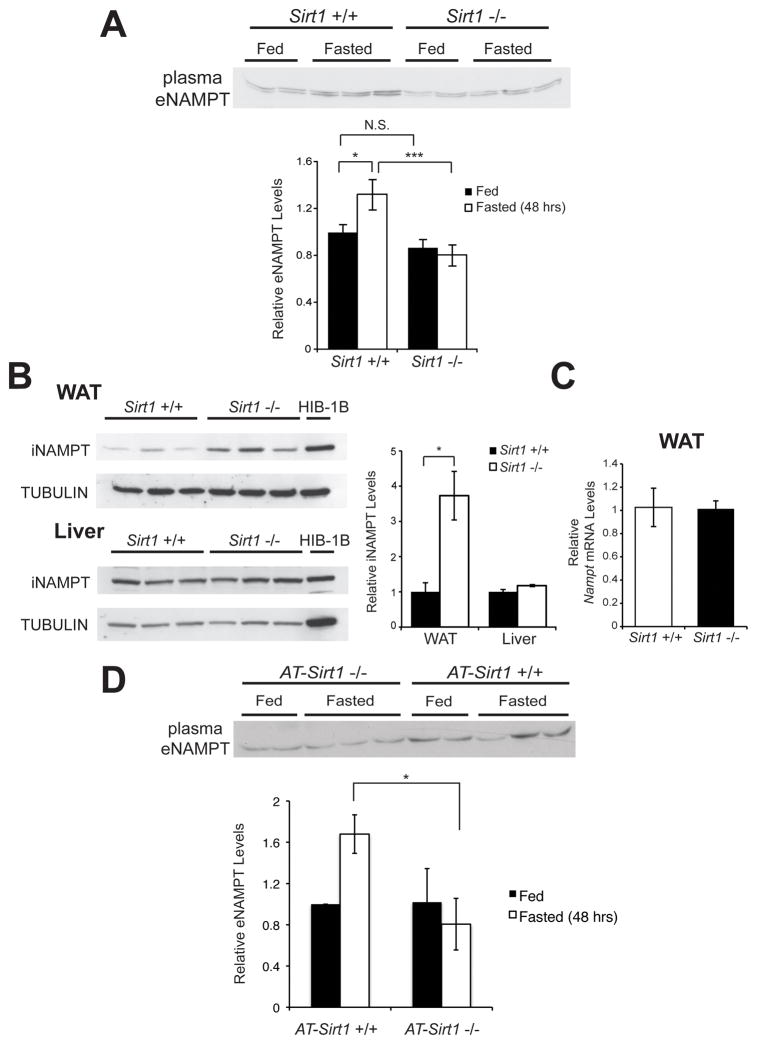
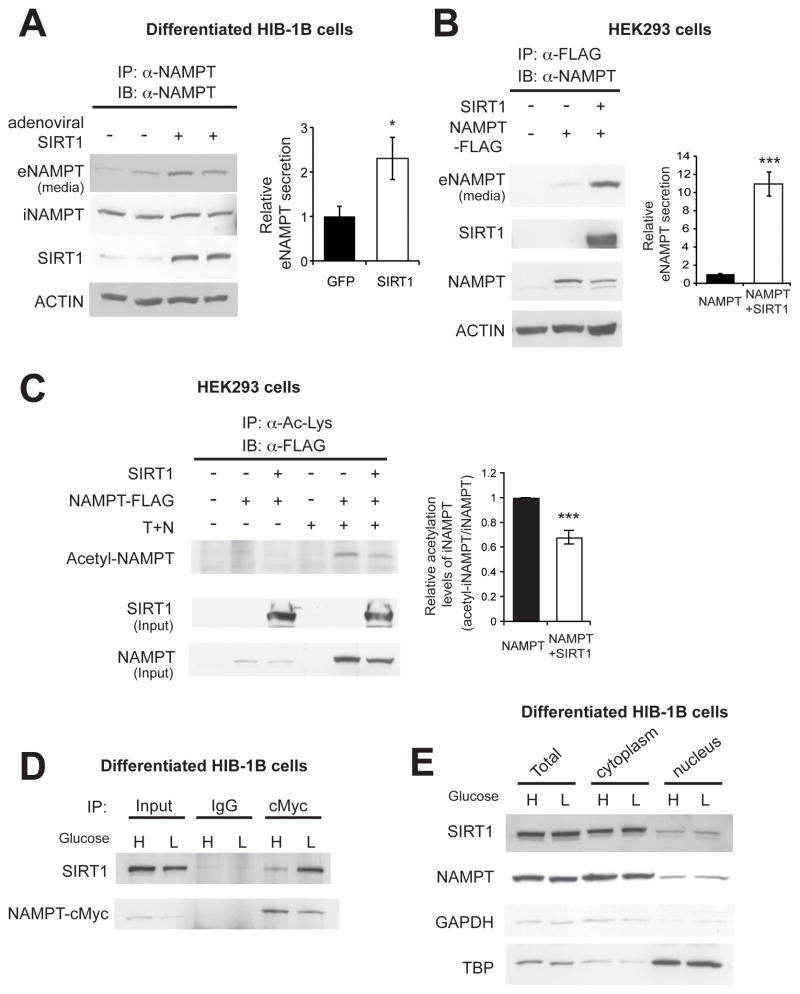
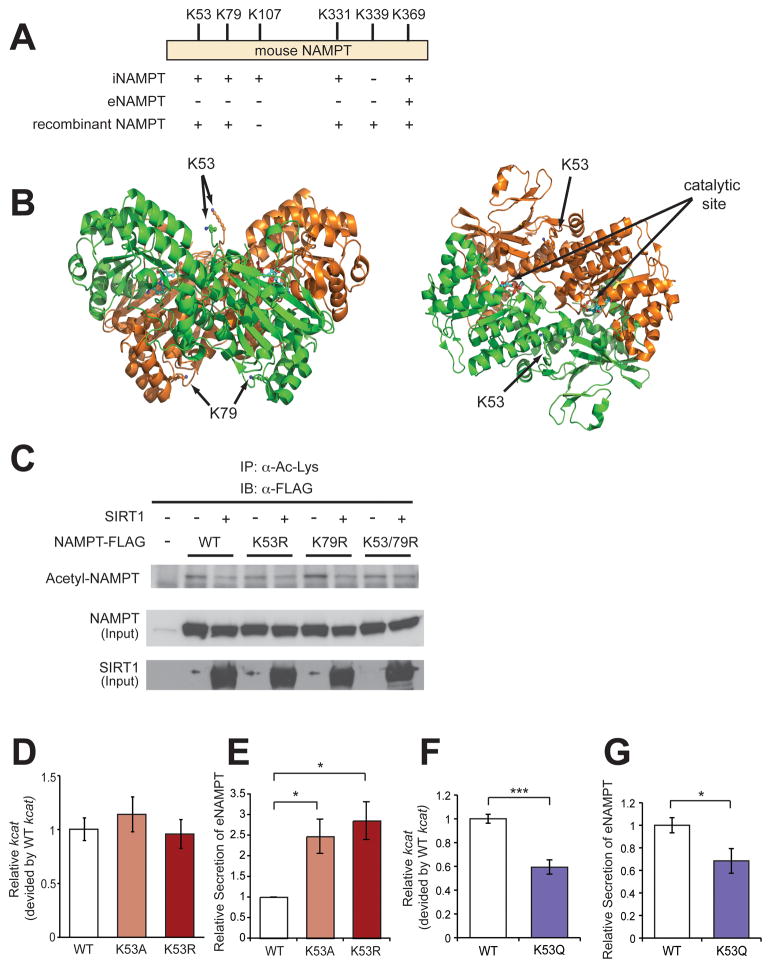
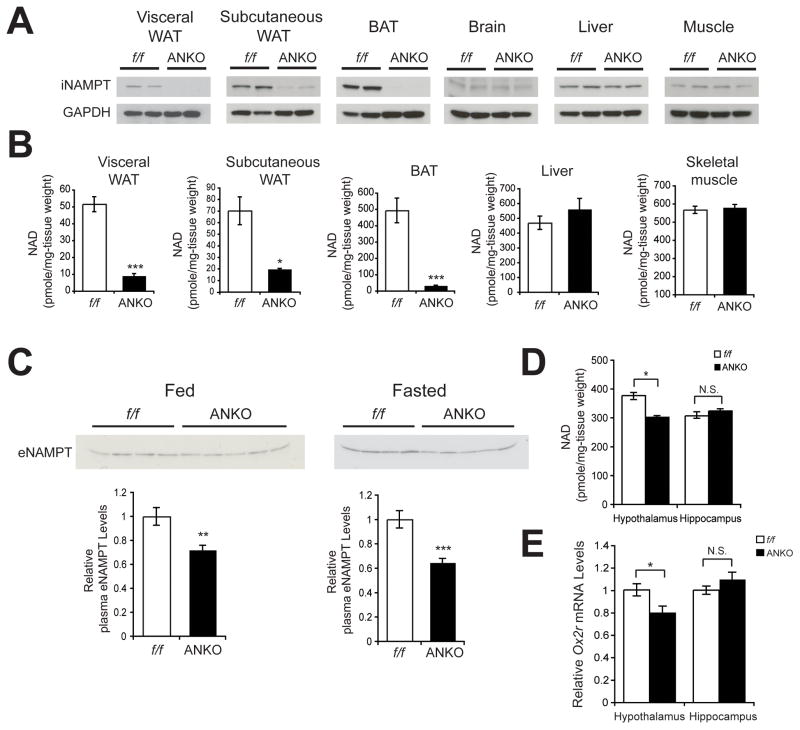
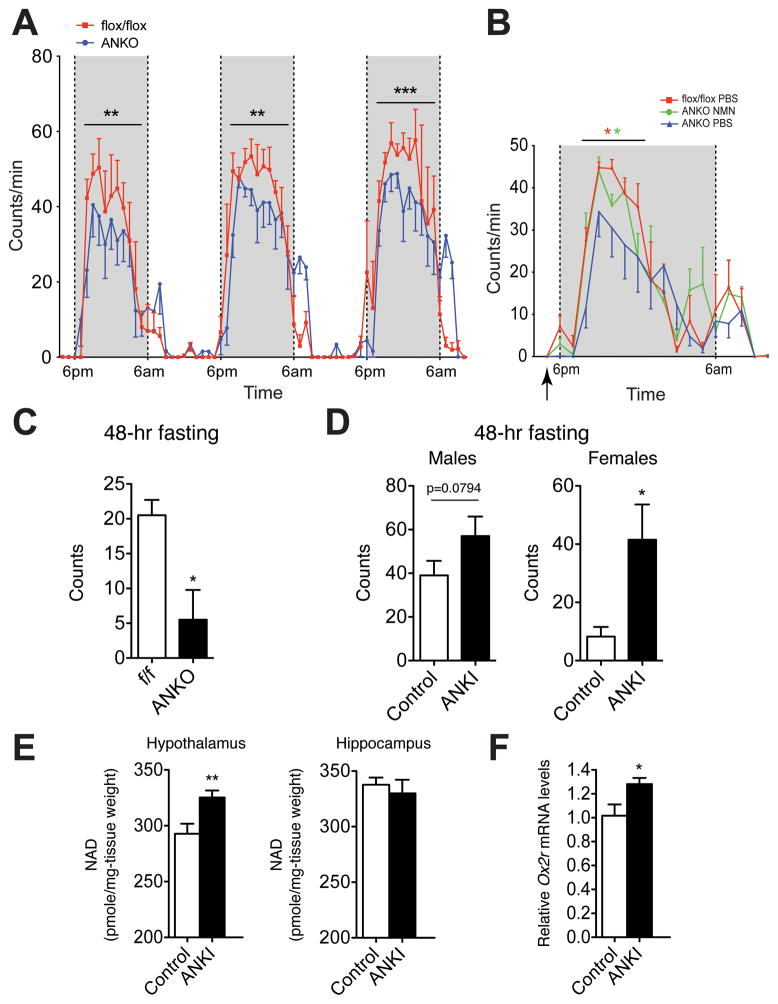
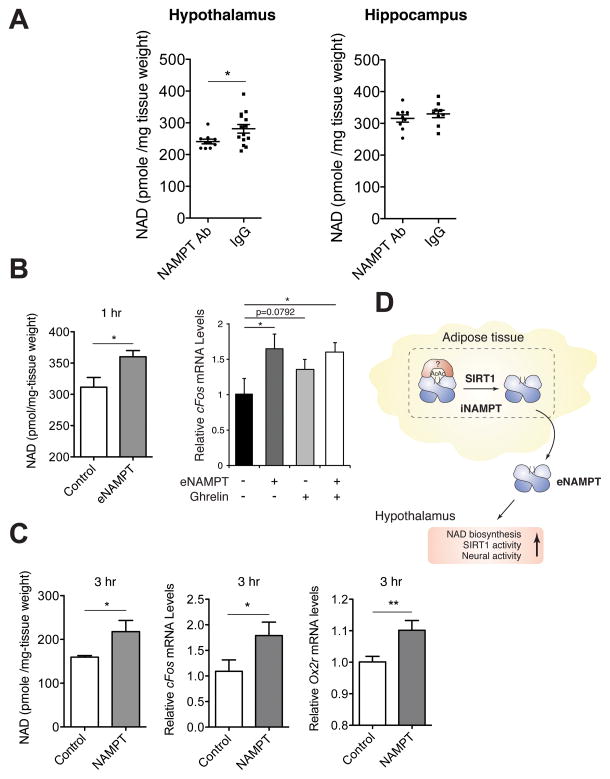
Nicotinamide phosphoribosyltransferase (NAMPT), the key NAD(+) biosynthetic enzyme, has two different forms, intra- and extracellular (iNAMPT and eNAMPT), in mammals. However, the significance of eNAMPT secretion remains unclear. Here we demonstrate that deacetylation of iNAMPT by the mammalian NAD(+)-dependent deacetylase SIRT1 predisposes the protein to secretion in adipocytes. NAMPT mutants reveal that SIRT1 deacetylates lysine 53 (K53) and enhances eNAMPT activity and secretion. Adipose tissue-specific Nampt knockout and knockin (ANKO and ANKI) mice show reciprocal changes in circulating eNAMPT, affecting hypothalamic NAD(+)/SIRT1 signaling and physical activity accordingly. The defect in physical activity observed in ANKO mice is ameliorated by nicotinamide mononucleotide (NMN). Furthermore, administration of a NAMPT-neutralizing antibody decreases hypothalamic NAD(+) production, and treating ex vivo hypothalamic explants with purified eNAMPT enhances NAD(+), SIRT1 activity, and neural activation. Thus, our findings indicate a critical role of adipose tissue as a modulator for the regulation of NAD(+) biosynthesis at a systemic level.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Bitterman KJ, Anderson RM, Cohen HY, Latorre-Esteves M, Sinclair DA. Inhibition of silencing and accelerated aging by nicotinamide, a putative negative regulator of yeast sir2 and human SIRT1. J Biol Chem. 2002;277:45099–45107. - PubMed
-
- Elmquist JK, Elias CF, Saper CB. From lesions to leptin: hypothalamic control of food intake and body weight. Neuron. 1999;22:221–232. - PubMed
-
- Fukuhara A, Matsuda M, Nishizawa M, Segawa K, Tanaka M, Kishimoto K, Matsuki Y, Murakami M, Ichisaka T, Murakami H, et al. Visfatin: a protein secreted by visceral fat that mimics the effects of insulin. Science. 2005;307:426–430. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
