

Use of both cumulus cells' transcriptomic markers and zona pellucida birefringence to select developmentally competent oocytes in human assisted reproductive technologies
- PMID: 25923296
- PMCID: PMC4315169
- DOI: 10.1186/1471-2164-16-S1-S9
Use of both cumulus cells' transcriptomic markers and zona pellucida birefringence to select developmentally competent oocytes in human assisted reproductive technologies
Abstract
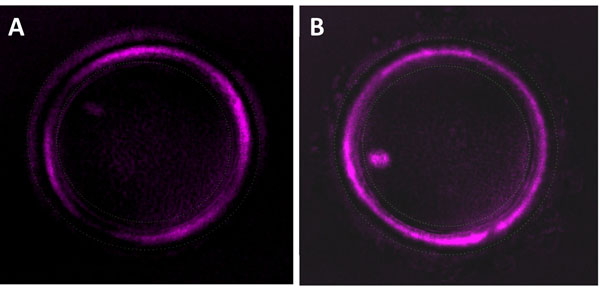
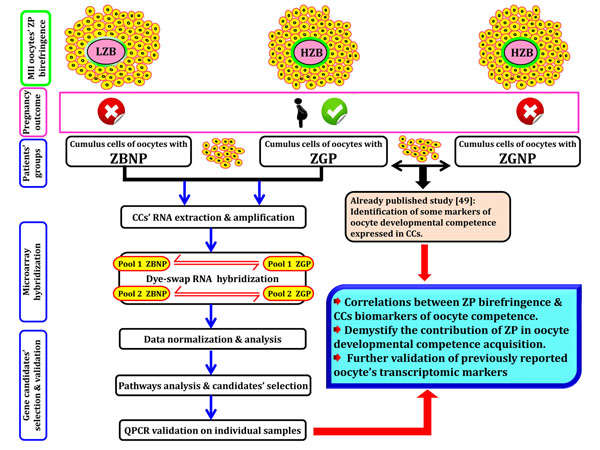
Background: Selection of the best oocyte for subsequent steps of fertilization and embryo transfer was shown to be the crucial step in human infertility treatment procedure. Oocyte selection using morphological criteria mainly Zona pellucida (ZP) has been the gold standard method in assisted reproductive technologies (ART) clinics, but this selection approach has limitations in terms of accuracy, objectivity and constancy. Recent studies using OMICs-based approaches have allowed the identification of key molecular markers that quantitatively and non-invasively predict the oocyte quality for higher pregnancy rates and efficient infertility treatment. These biomarkers are a valuable reinforcement of the morphological selection criteria widely used in in vitro fertilization (IVF) clinics. In this context, this study was designed to investigate the relationship between transcriptomic predictors of oocyte quality found by our group and the conventional morphological parameters of oocyte quality mainly the ZP birefringence.
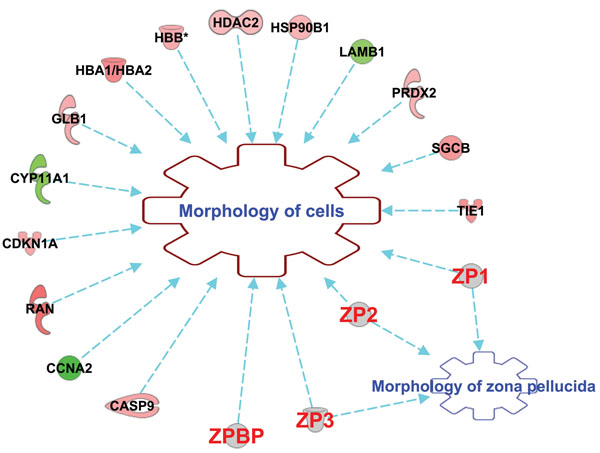
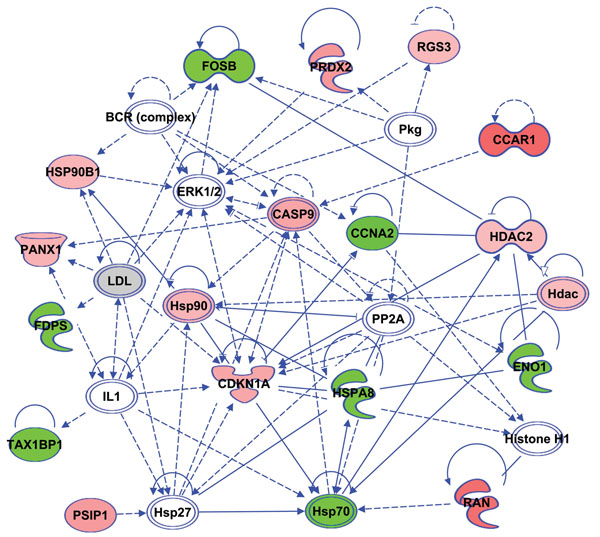
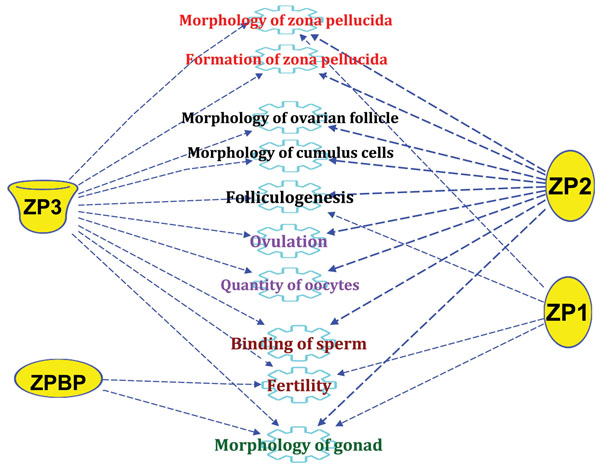
Results: Microarray data revealed that 48 and 27 differentially expressed candidate genes in cumulus cells (CCs) were respectively overexpressed and underexpressed in the ZGP (Zona Good Pregnant) versus ZBNP (Zona Bad Non Pregnant) groups. More than 70% of previously reported transcriptomic biomarkers of oocyte developmental competence were confirmed in this study. The analysis of possible association between ZP birefringence versus molecular markers approach showed an absence of correlation between them using the current set of markers.
Conclusions: This study suggested a new integrative approach that matches morphological and molecular approaches used to select developmentally competent oocytes able to lead to successful pregnancy and the delivery of healthy baby. For each ZP birefringence score, oocytes displayed a particular CCs' gene expression pattern. However, no correlations were found between the 7 gene biomarkers of oocyte developmental potential and the ZP birefringence score. Further studies using larger lists of candidate markers are required to identify suitable genes that are highly correlated with the morphological criteria, and therefore able to reinforce the accuracy of oocyte selection and the effectiveness of infertility treatment.
Figures


References
-
- Zegers-Hochschild F, Adamson GD, de Mouzon J, Ishihara O, Mansour R, Nygren K, Sullivan E, Vanderpoel S. International Committee for Monitoring Assisted Reproductive Technology (ICMART) and the World Health Organization (WHO) revised glossary of ART terminology, 2009. Fertility and sterility. 2009;92(5):1520–1524. doi: 10.1016/j.fertnstert.2009.09.009. - DOI - PubMed
-
- Krisher RL. The effect of oocyte quality on development. J Anim Sci. 2004;82(E-Suppl):E14–23. - PubMed
-
- Sirard M-A, Assidi M. In: Oocyte Physiology and Development in Domestic Animals. Krisher RL, editor. Wiley-Blackwell; 2013. Screening for Oocyte Competence; pp. 191–206.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
