

Translation initiation factor eIF3 promotes programmed stop codon readthrough
- PMID: 25925566
- PMCID: PMC4446449
- DOI: 10.1093/nar/gkv421
Translation initiation factor eIF3 promotes programmed stop codon readthrough
Abstract
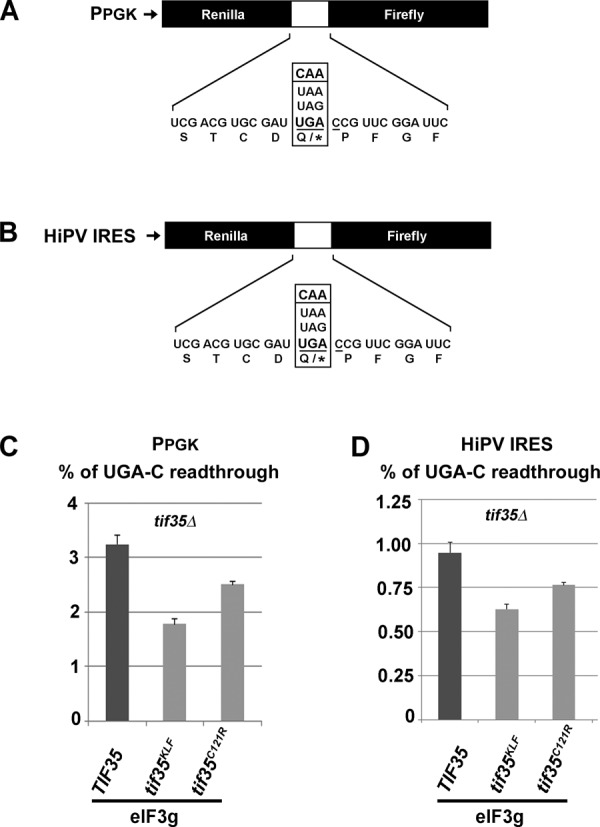
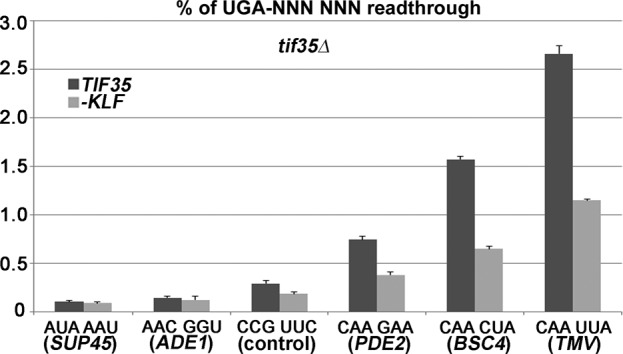
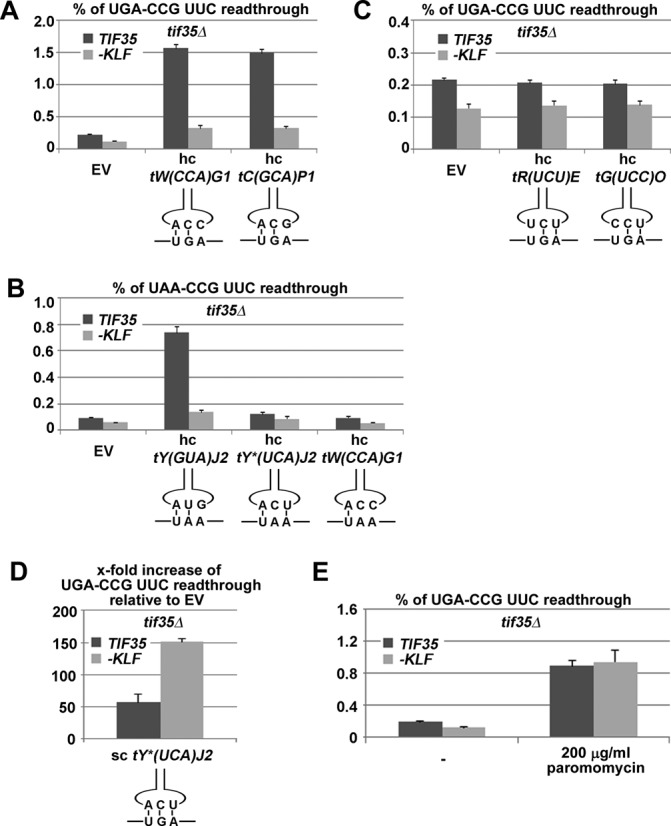
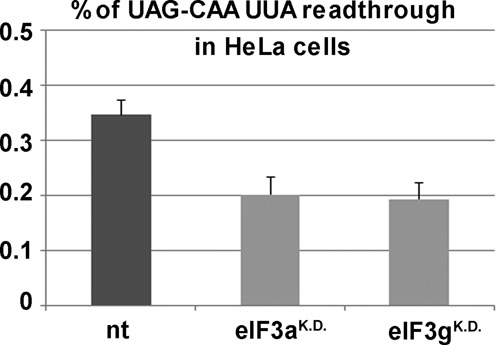
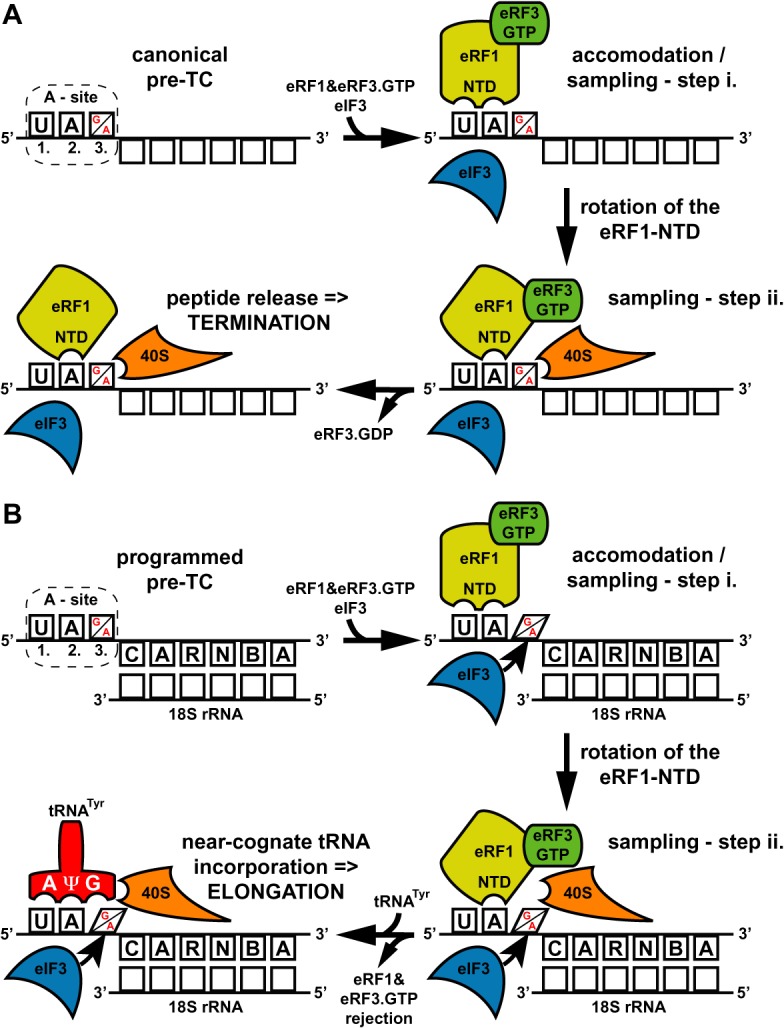
Programmed stop codon readthrough is a post-transcription regulatory mechanism specifically increasing proteome diversity by creating a pool of C-terminally extended proteins. During this process, the stop codon is decoded as a sense codon by a near-cognate tRNA, which programs the ribosome to continue elongation. The efficiency of competition for the stop codon between release factors (eRFs) and near-cognate tRNAs is largely dependent on its nucleotide context; however, the molecular mechanism underlying this process is unknown. Here, we show that it is the translation initiation (not termination) factor, namely eIF3, which critically promotes programmed readthrough on all three stop codons. In order to do so, eIF3 must associate with pre-termination complexes where it interferes with the eRF1 decoding of the third/wobble position of the stop codon set in the unfavorable termination context, thus allowing incorporation of near-cognate tRNAs with a mismatch at the same position. We clearly demonstrate that efficient readthrough is enabled by near-cognate tRNAs with a mismatch only at the third/wobble position. Importantly, the eIF3 role in programmed readthrough is conserved between yeast and humans.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures


References
-
- Jackson R.J., Hellen C.U., Pestova T.V. Termination and post-termination events in eukaryotic translation. Adv. Protein Chem. Struct. Biol. 2012;86:45–93. - PubMed
-
- Robinson D.N., Cooley L. Examination of the function of two kelch proteins generated by stop codon suppression. Development. 1997;124:1405–1417. - PubMed
-
- Bonetti B., Fu L., Moon J., Bedwell D.M. The efficiency of translation termination is determined by a synergistic interplay between upstream and downstream sequences in Saccharomyces cerevisiae. J. Mol. Biol. 1995;251:334–345. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases

