

The transmembrane protein Crumbs displays complex dynamics during follicular morphogenesis and is regulated competitively by Moesin and aPKC
- PMID: 25926360
- PMCID: PMC4440922
- DOI: 10.1242/dev.115329
The transmembrane protein Crumbs displays complex dynamics during follicular morphogenesis and is regulated competitively by Moesin and aPKC
Erratum in
-
The transmembrane protein Crumbs displays complex dynamics during follicular morphogenesis and is regulated competitively by Moesin and aPKC.Development. 2015 Jun 15;142(12):2226. doi: 10.1242/dev.126425. Development. 2015. PMID: 26081574 Free PMC article. No abstract available.
Abstract
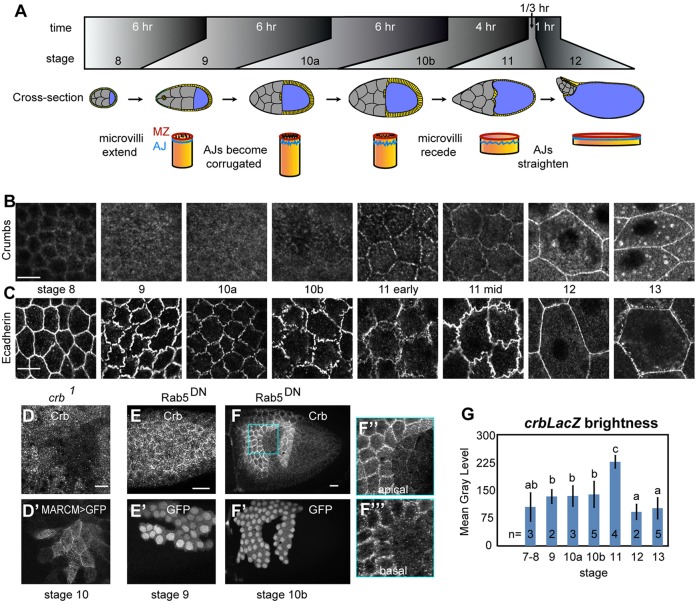
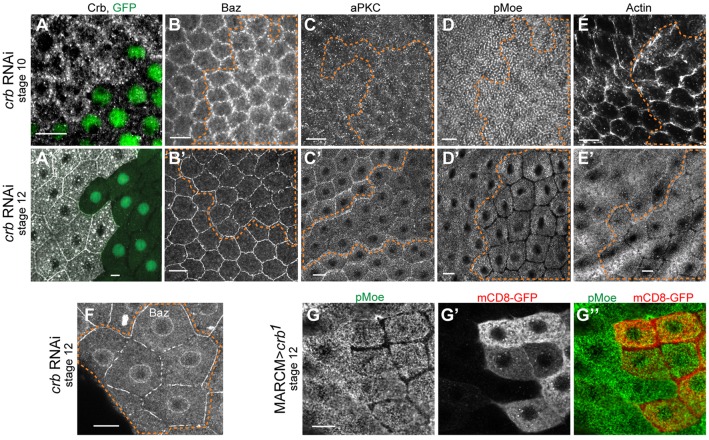
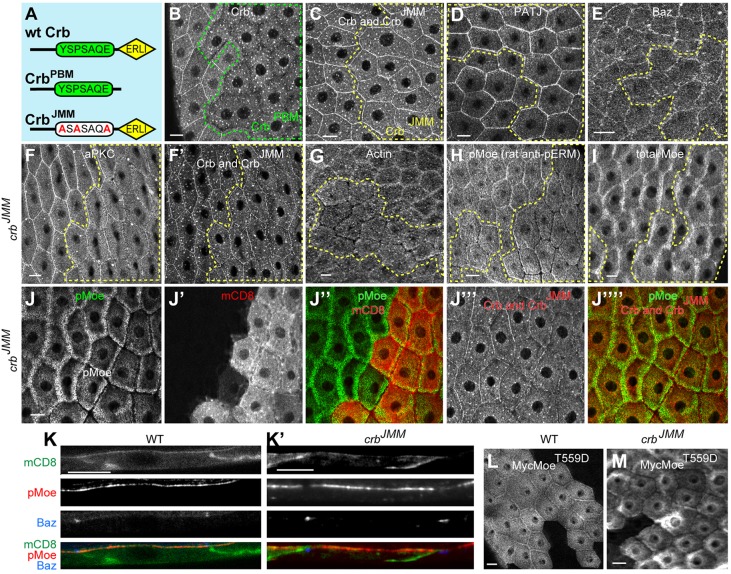
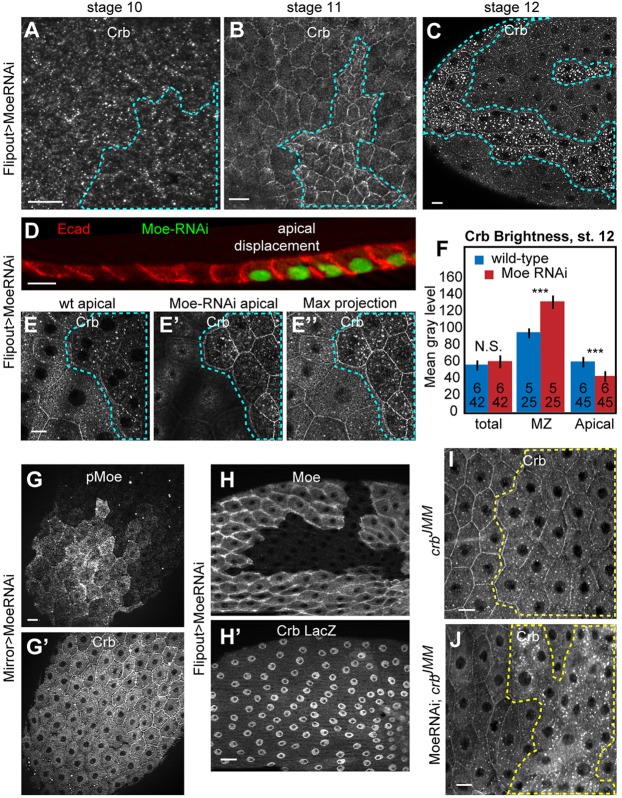
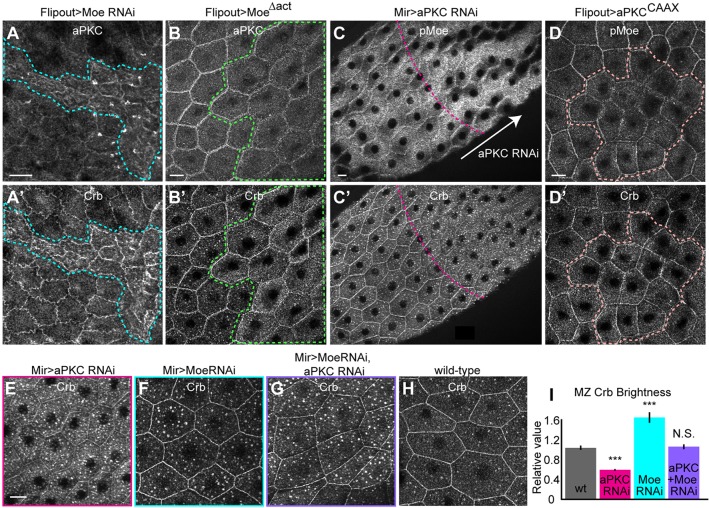
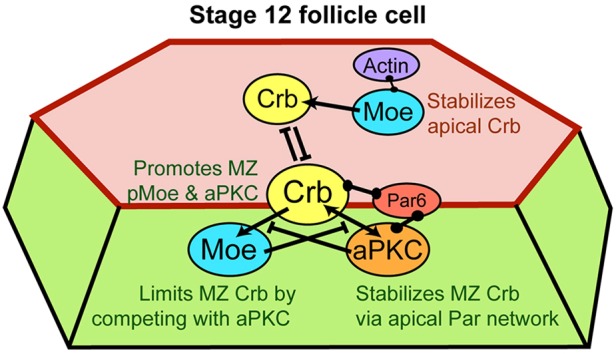
The transmembrane protein Crumbs (Crb) functions in apical polarity and epithelial integrity. To better understand its role in epithelial morphogenesis, we examined Crb localization and dynamics in the late follicular epithelium of Drosophila. Crb was unexpectedly dynamic during middle-to-late stages of egg chamber development, being lost from the marginal zone (MZ) in stage 9 before abruptly returning at the end of stage 10b, then undergoing a pulse of endocytosis in stage 12. The reappearance of MZ Crb is necessary to maintain an intact adherens junction and MZ. Although Crb has been proposed to interact through its juxtamembrane domain with Moesin (Moe), a FERM domain protein that regulates the cortical actin cytoskeleton, the functional significance of this interaction is poorly understood. We found that whereas the Crb juxtamembrane domain was not required for adherens junction integrity, it was necessary for MZ localization of Moe, aPKC and F-actin. Furthermore, Moe and aPKC functioned antagonistically, suggesting that Moe limits Crb levels by reducing its interactions with the apical Par network. Additionally, Moe mutant cells lost Crb from the apical membrane and accumulated excess Crb at the MZ, suggesting that Moe regulates Crb distribution at the membrane. Together, these studies reveal reciprocal interactions between Crb, Moe and aPKC during cellular morphogenesis.
Keywords: Crumbs; ERM proteins; F-actin; Par3/Bazooka; Squamous morphogenesis; aPKC.
© 2015. Published by The Company of Biologists Ltd.
Figures
Similar articles
-
Cdc42 defines apical identity and regulates epithelial morphogenesis by promoting apical recruitment of Par6-aPKC and Crumbs.Development. 2019 Aug 12;146(15):dev175497. doi: 10.1242/dev.175497. Development. 2019. PMID: 31405903 Free PMC article.
-
Structural basis for the phosphorylation-regulated interaction between the cytoplasmic tail of cell polarity protein crumbs and the actin-binding protein moesin.J Biol Chem. 2015 May 1;290(18):11384-92. doi: 10.1074/jbc.M115.643791. Epub 2015 Mar 19. J Biol Chem. 2015. PMID: 25792740 Free PMC article.
-
Regulated Crb accumulation controls apical constriction and invagination in Drosophila tracheal cells.J Cell Sci. 2011 Jan 15;124(Pt 2):240-51. doi: 10.1242/jcs.073601. Epub 2010 Dec 15. J Cell Sci. 2011. PMID: 21172808
-
PAR-Complex and Crumbs Function During Photoreceptor Morphogenesis and Retinal Degeneration.Front Cell Neurosci. 2018 Mar 29;12:90. doi: 10.3389/fncel.2018.00090. eCollection 2018. Front Cell Neurosci. 2018. PMID: 29651238 Free PMC article. Review.
-
The apical polarity protein network in Drosophila epithelial cells: regulation of polarity, junctions, morphogenesis, cell growth, and survival.Annu Rev Cell Dev Biol. 2012;28:655-85. doi: 10.1146/annurev-cellbio-092910-154033. Epub 2012 Aug 6. Annu Rev Cell Dev Biol. 2012. PMID: 22881460 Review.
Cited by
-
Crumbs, Moesin and Yurt regulate junctional stability and dynamics for a proper morphogenesis of the Drosophila pupal wing epithelium.Sci Rep. 2017 Dec 1;7(1):16778. doi: 10.1038/s41598-017-15272-1. Sci Rep. 2017. PMID: 29196707 Free PMC article.
-
Using Drosophila Nephrocytes to Understand the Formation and Maintenance of the Podocyte Slit Diaphragm.Front Cell Dev Biol. 2022 Feb 21;10:837828. doi: 10.3389/fcell.2022.837828. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35265622 Free PMC article. Review.
-
Distinct functions of Crumbs regulating slit diaphragms and endocytosis in Drosophila nephrocytes.Cell Mol Life Sci. 2017 Dec;74(24):4573-4586. doi: 10.1007/s00018-017-2593-y. Epub 2017 Jul 17. Cell Mol Life Sci. 2017. PMID: 28717874 Free PMC article.
-
Comparing statistical learning methods for complex trait prediction from gene expression.PLoS One. 2025 Feb 11;20(2):e0317516. doi: 10.1371/journal.pone.0317516. eCollection 2025. PLoS One. 2025. PMID: 39932918 Free PMC article.
-
Comparing statistical learning methods for complex trait prediction from gene expression.bioRxiv [Preprint]. 2024 Jun 3:2024.06.01.596951. doi: 10.1101/2024.06.01.596951. bioRxiv. 2024. Update in: PLoS One. 2025 Feb 11;20(2):e0317516. doi: 10.1371/journal.pone.0317516. PMID: 38895364 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
