

Human Endogenous Retrovirus Type K (HERV-K) Particles Package and Transmit HERV-K-Related Sequences
- PMID: 25926654
- PMCID: PMC4473553
- DOI: 10.1128/JVI.00544-15
Human Endogenous Retrovirus Type K (HERV-K) Particles Package and Transmit HERV-K-Related Sequences
Abstract
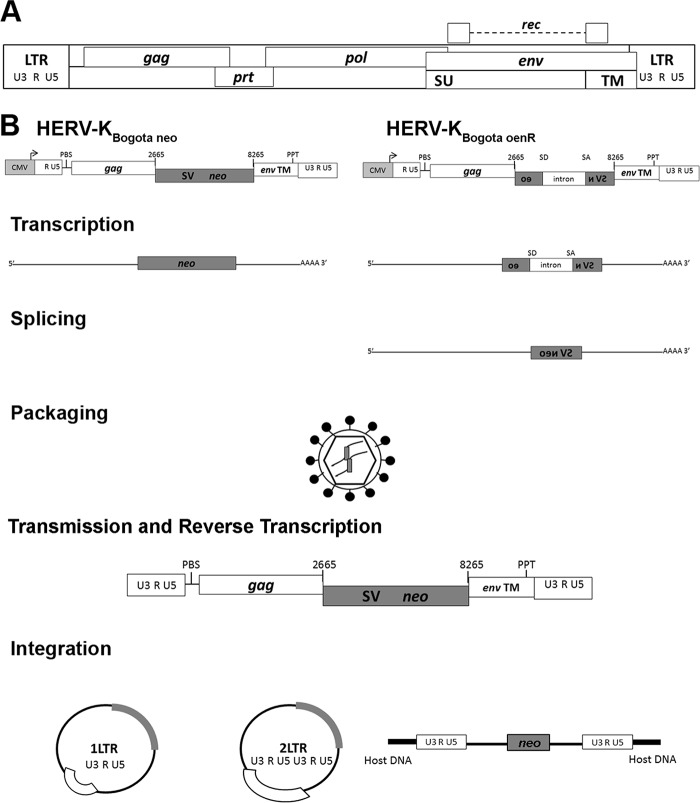
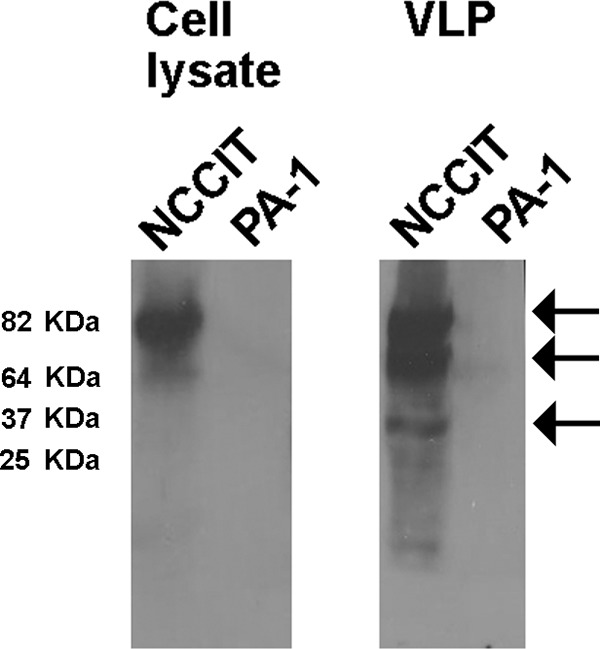
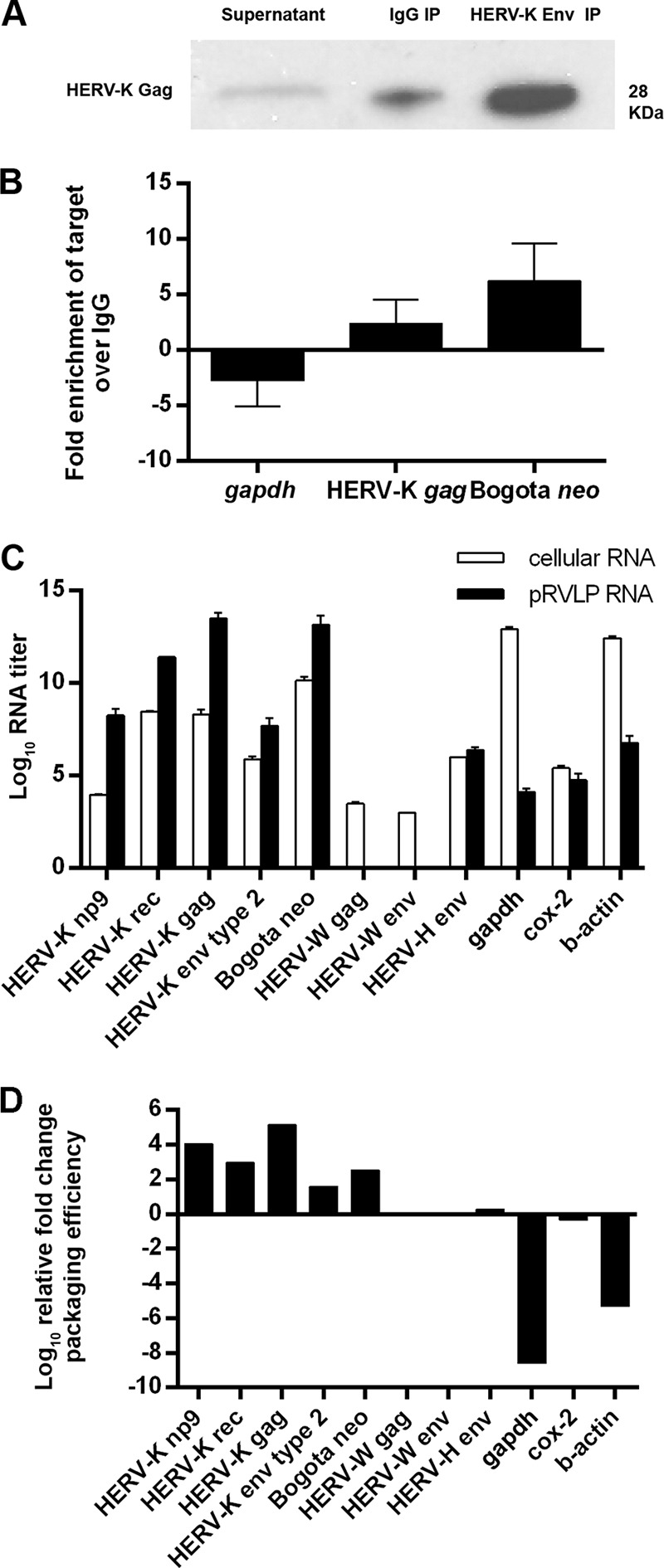
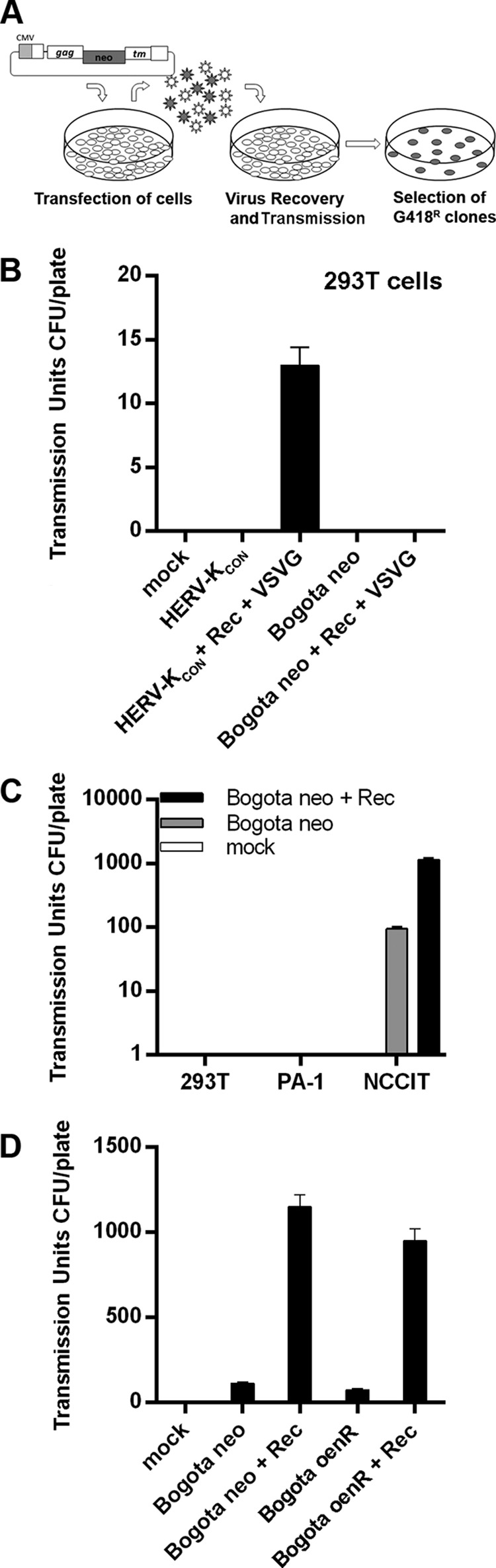
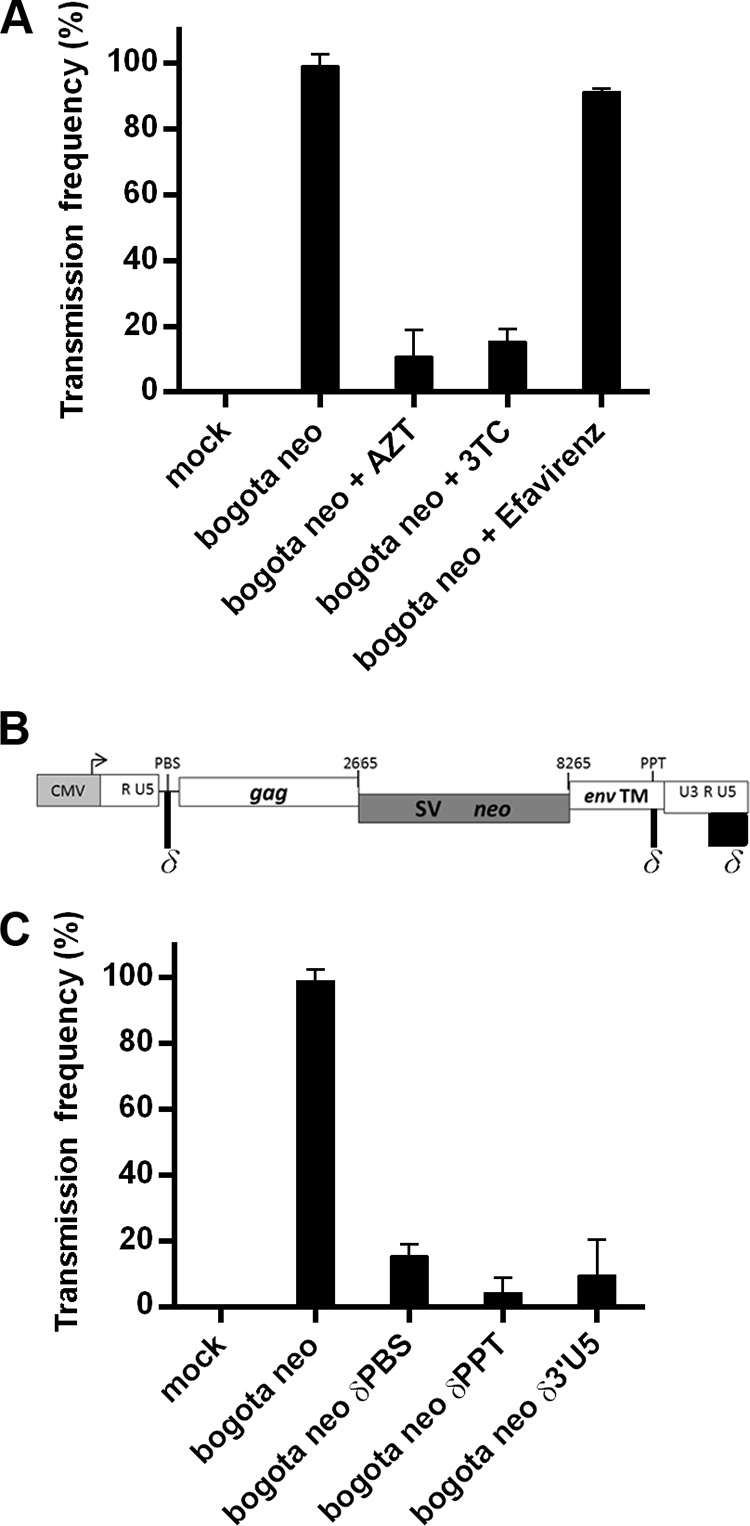
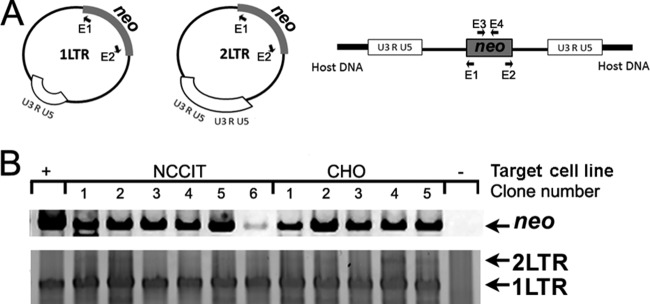
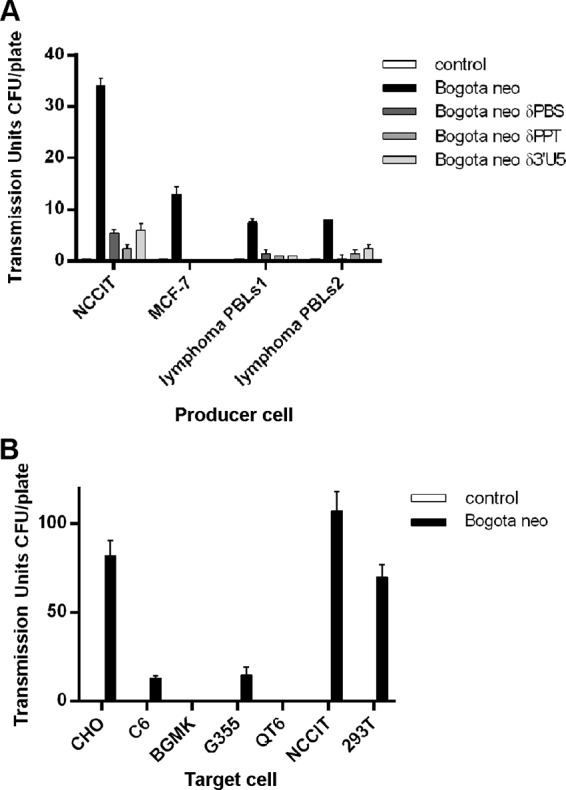
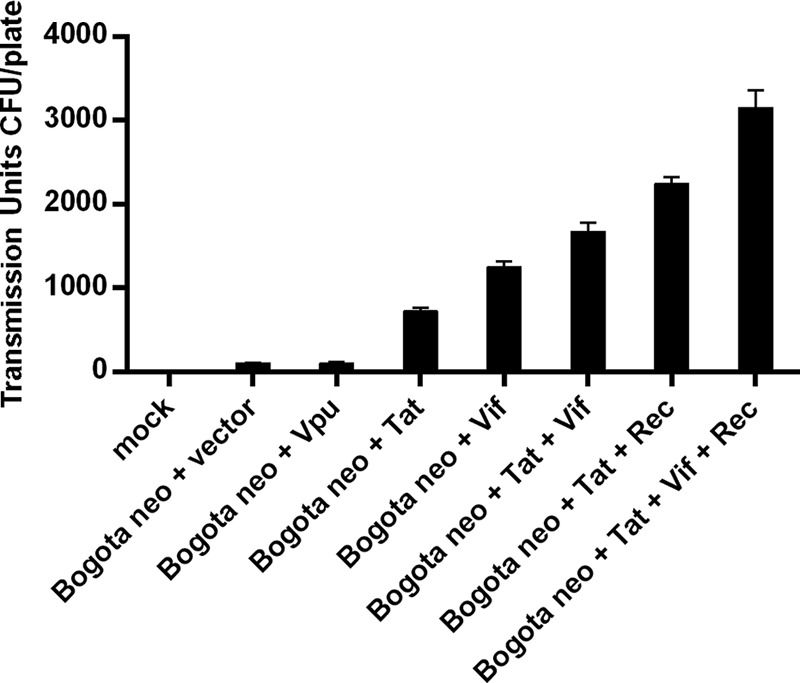
Human endogenous retroviruses (HERV) make up 8% of the human genome. While the youngest of these retroviruses, HERV-K(HML-2), termed HK2, is able to code for all viral proteins and produce virus-like particles, it is not known if these virus particles package and transmit HK2-related sequences. Here, we analyzed the capacity of HK2 for packaging and transmitting HK2 sequences. We created an HK2 probe, termed Bogota, which can be packaged into HK2 viruses, and transfected it into cells that make HK2 particles. Supernatants of the transfected cells, which contained HK2 viral particles, then were added to target cells, and the transmissibility of the HK2 Bogota reporter was tracked by G418 resistance. Our studies revealed that contemporary HK2 virions produced by some teratocarcinoma and breast cancer cell lines, as well as by peripheral blood lymphocytes from lymphoma patients, can package HK2 Bogota probes, and these viruses transmitted these probes to other cells. After transmission, HK2 Bogota transcripts undergo reverse transcription, a step impaired by antiretroviral agents or by introduction of mutations into the probe sequences required for reverse transcription. HK2 viruses were more efficiently transmitted in the presence of HK2 Rec or HIV-1 Tat and Vif. Transmitted Bogota probes formed episomes but did not integrate into the cellular genome. Resistance to integration might explain the relatively low number of HK2 insertions that were acquired during the last 25 million years of evolution. Whether transient transmission of modern HK2 sequences, which encode two putative oncoproteins, can lead to disease remains to be studied.
Importance: Retroviruses invaded the genome of human ancestors over the course of millions of years, yet these viruses generally have been inactivated during evolution, with only remnants of these infectious sequences remaining in the human genome. One of these viruses, termed HK2, still is capable of producing virus particles, although these particles have been regarded as being noninfectious. Using a genetic probe derived from HK2, we have discovered that HK2 viruses produced in modern humans can package HK2 sequences and transmit them to various other cells. Furthermore, the genetic sequences packaged in HK2 undergo reverse transcription. The transmitted probe circularized in the cell and failed to integrate into the cellular genome. These findings suggest that modern HK2 viruses can package viral RNA and transmit it to other cells. Contrary to previous views, we provide evidence of an extracellular viral phase of modern HK2 viruses. We have no evidence of sustained, spreading infection.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- T32 AI007528/AI/NIAID NIH HHS/United States
- 1F31CA150523-01/CA/NCI NIH HHS/United States
- 5T32AI007528-13/AI/NIAID NIH HHS/United States
- T32 GM145304/GM/NIGMS NIH HHS/United States
- F31 CA150523/CA/NCI NIH HHS/United States
- 3R01CA144043-03S1/CA/NCI NIH HHS/United States
- U54 DA021519/DA/NIDA NIH HHS/United States
- UL1RR24986/RR/NCRR NIH HHS/United States
- RM-08-029/RM/RMOD NIH HHS/United States
- U54DA021519/DA/NIDA NIH HHS/United States
- K22 CA177824/CA/NCI NIH HHS/United States
- F32 AI106189/AI/NIAID NIH HHS/United States
- P30 ES017885/ES/NIEHS NIH HHS/United States
- UL1 RR024986/RR/NCRR NIH HHS/United States
- P30U54ES017885/ES/NIEHS NIH HHS/United States
- R01 CA144043/CA/NCI NIH HHS/United States
- T32 GM007315/GM/NIGMS NIH HHS/United States
- 1 F32 AI106189-01/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
