

Activation and regulation of DNA-driven immune responses
- PMID: 25926682
- PMCID: PMC4429241
- DOI: 10.1128/MMBR.00061-14
Activation and regulation of DNA-driven immune responses
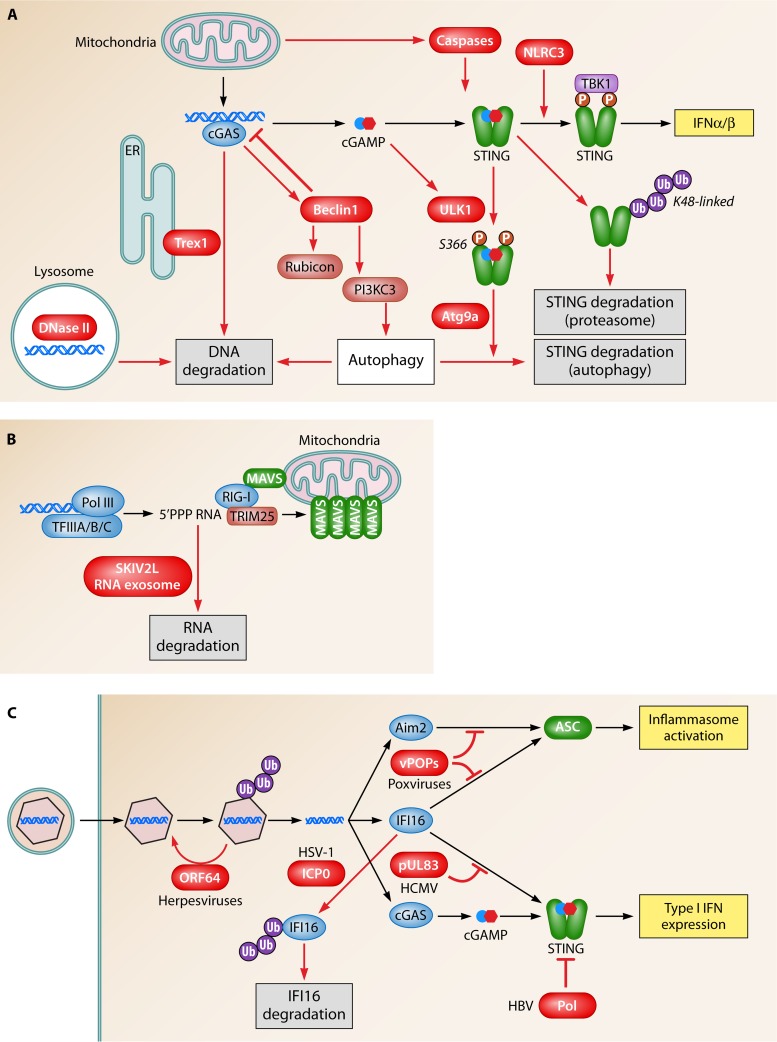
Abstract
The innate immune system provides early defense against infections and also plays a key role in monitoring alterations of homeostasis in the body. DNA is highly immunostimulatory, and recent advances in this field have led to the identification of the innate immune sensors responsible for the recognition of DNA as well as the downstream pathways that are activated. Moreover, information on how cells regulate DNA-driven immune responses to avoid excessive inflammation is now emerging. Finally, several reports have demonstrated how defects in DNA sensing, signaling, and regulation are associated with susceptibility to infections or inflammatory diseases in humans and model organisms. In this review, the current literature on DNA-stimulated innate immune activation is discussed, and important new questions facing this field are proposed.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
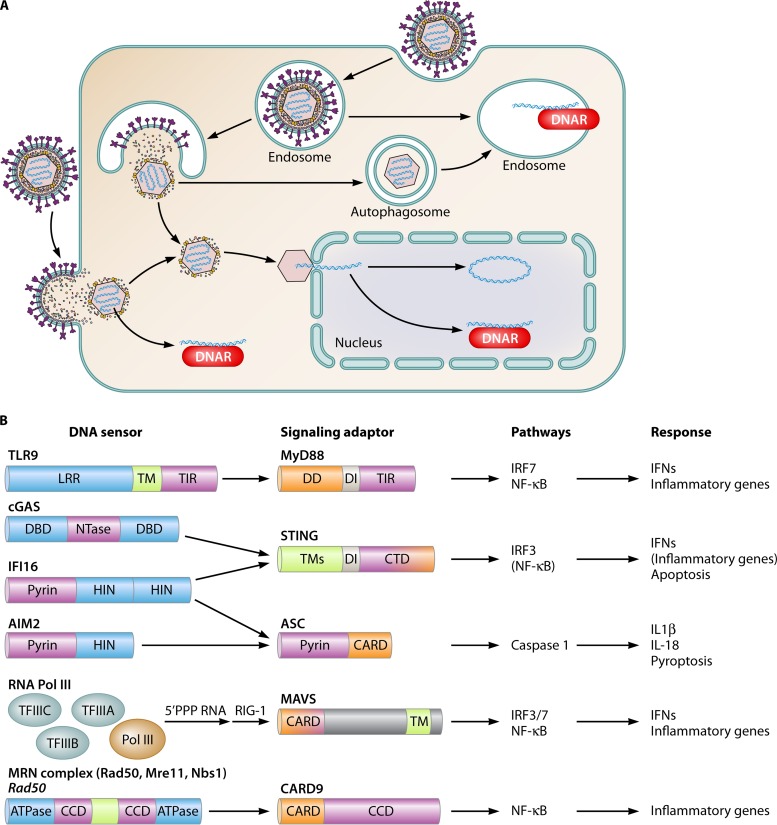
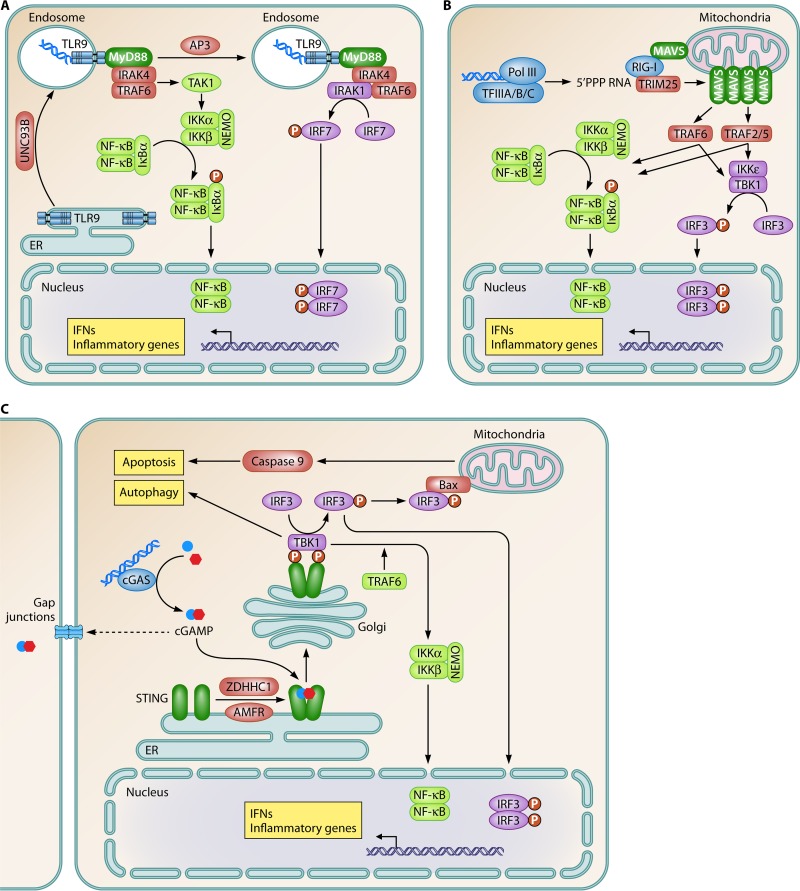
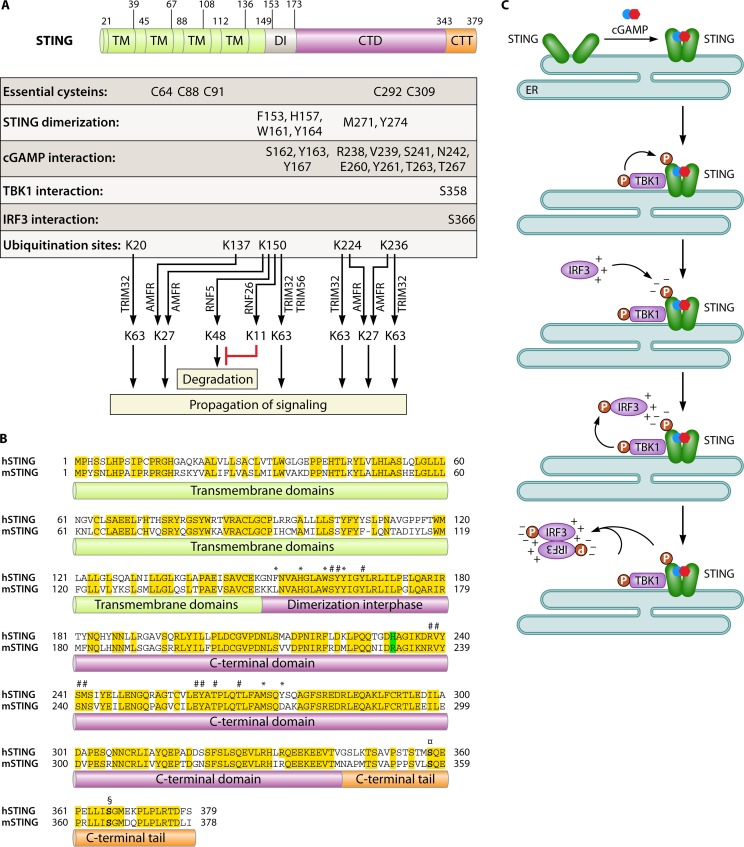
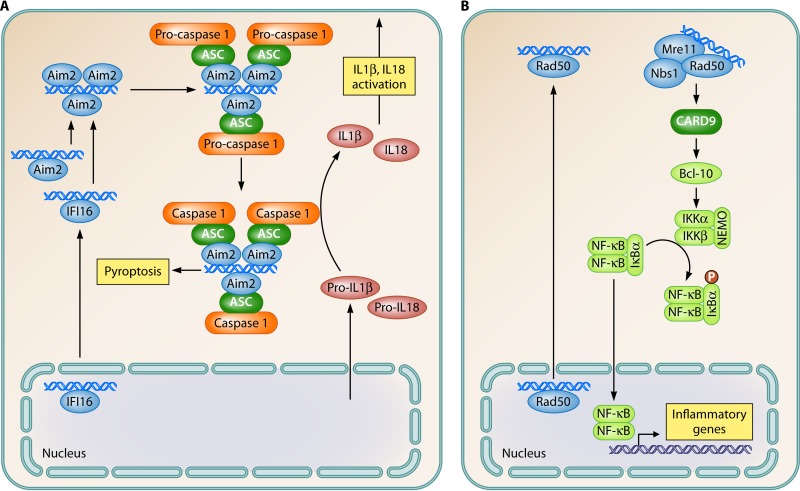
References
-
- Ishii KJ, Coban C, Kato H, Takahashi K, Torii Y, Takeshita F, Ludwig H, Sutter G, Suzuki K, Hemmi H, Sato S, Yamamoto M, Uematsu S, Kawai T, Takeuchi O, Akira S. 2006. A Toll-like receptor-independent antiviral response induced by double-stranded B-form DNA. Nat Immunol 7:40–48. doi: 10.1038/ni1282. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
