

Negative Regulation of Humoral Immunity Due to Interplay between the SLAMF1, SLAMF5, and SLAMF6 Receptors
- PMID: 25926831
- PMCID: PMC4396446
- DOI: 10.3389/fimmu.2015.00158
Negative Regulation of Humoral Immunity Due to Interplay between the SLAMF1, SLAMF5, and SLAMF6 Receptors
Abstract
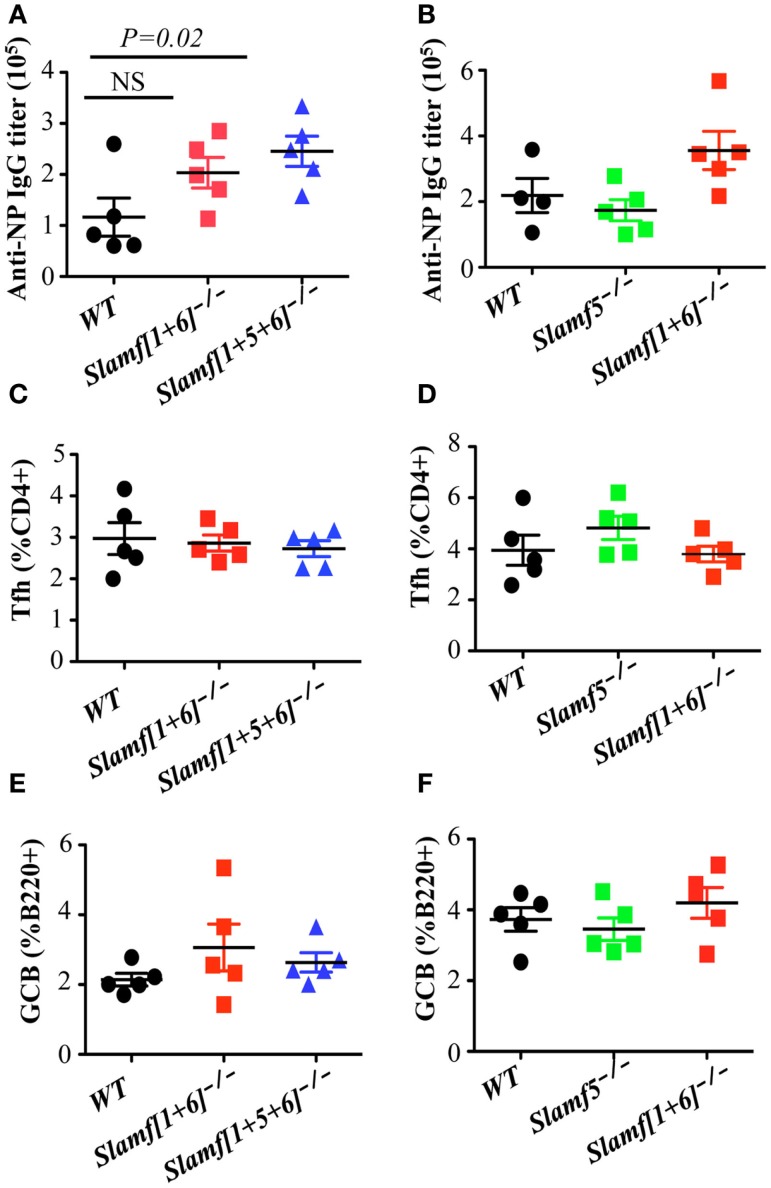
Whereas the SLAMF-associated protein (SAP) is involved in differentiation of T follicular helper (Tfh) cells and antibody responses, the precise requirements of SLAMF receptors in humoral immune responses are incompletely understood. By analyzing mice with targeted disruptions of the Slamf1, Slamf5, and Slamf6 genes, we found that both T-dependent and T-independent antibody responses were twofold higher compared to those in single knockout mice. These data suggest a suppressive synergy of SLAMF1, SLAMF5, and SLAMF6 in humoral immunity, which contrasts the decreased antibody responses resulting from a defective GC reaction in the absence of the adapter SAP. In adoptive co-transfer assays, both [Slamf1 + 5 + 6] (-/-) B and T cells were capable of inducing enhanced antibody responses, but more pronounced enhancement was observed after adoptive transfer of [Slamf1 + 5 + 6] (-/-) B cells compared to that of [Slamf1 + 5 + 6] (-/-) T cells. In support of [Slamf1 + 5 + 6] (-/-) B cell intrinsic activity, [Slamf1 + 5 + 6] (-/-) mice also mounted significantly higher antibody responses to T-independent type 2 antigen. Furthermore, treatment of mice with anti-SLAMF6 monoclonal antibody results in severe inhibition of the development of Tfh cells and GC B cells, confirming a suppressive effect of SLAMF6. Taken together, these results establish SLAMF1, SLAMF5, and SLAMF6 as important negative regulators of humoral immune response, consistent with the notion that SLAM family receptors have dual functions in immune responses.
Keywords: SLAM family receptors; SLAM-associated protein; anti-SLAMF6 mAb; follicular T helper cells; germinal center B cells; marginal zone B cells.
Figures










References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous