

Nanoscale Synaptic Membrane Mimetic Allows Unbiased High Throughput Screen That Targets Binding Sites for Alzheimer's-Associated Aβ Oligomers
- PMID: 25928376
- PMCID: PMC4415972
- DOI: 10.1371/journal.pone.0125263
Nanoscale Synaptic Membrane Mimetic Allows Unbiased High Throughput Screen That Targets Binding Sites for Alzheimer's-Associated Aβ Oligomers
Abstract
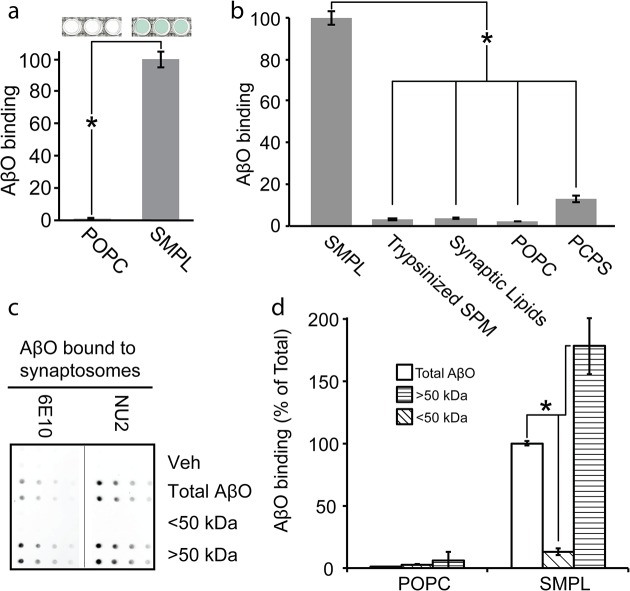
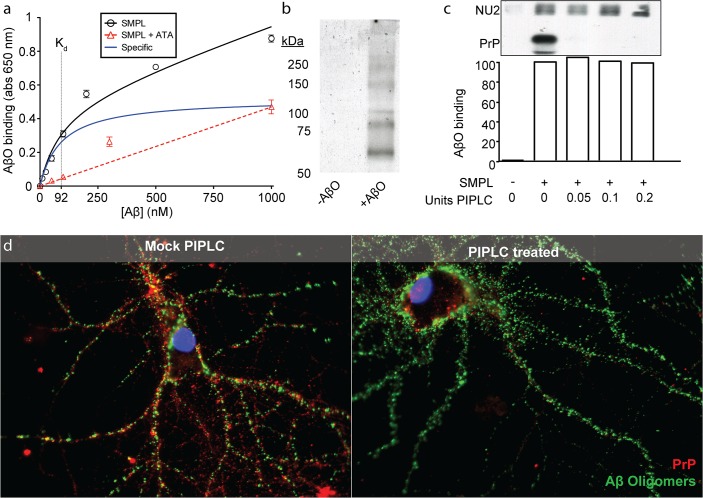
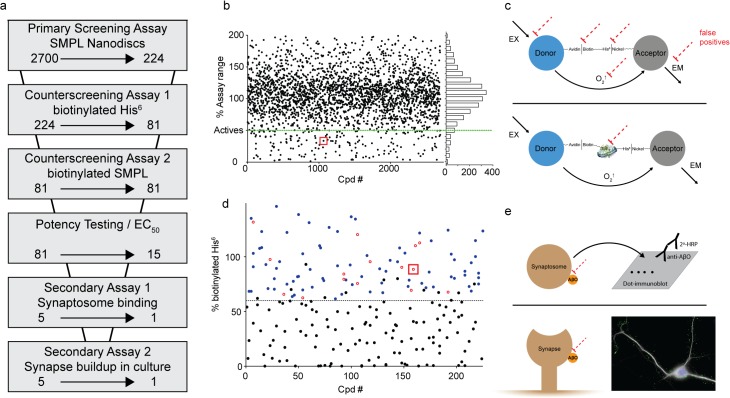
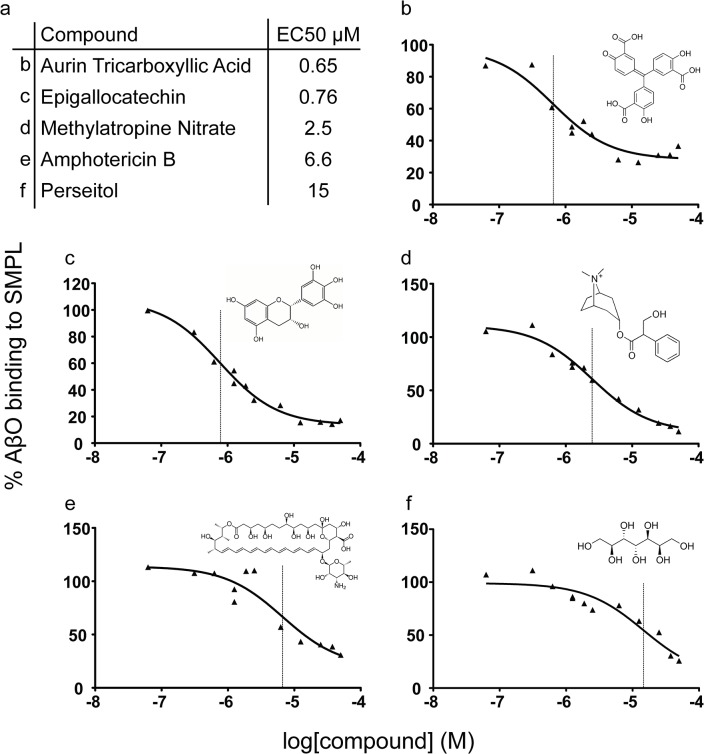
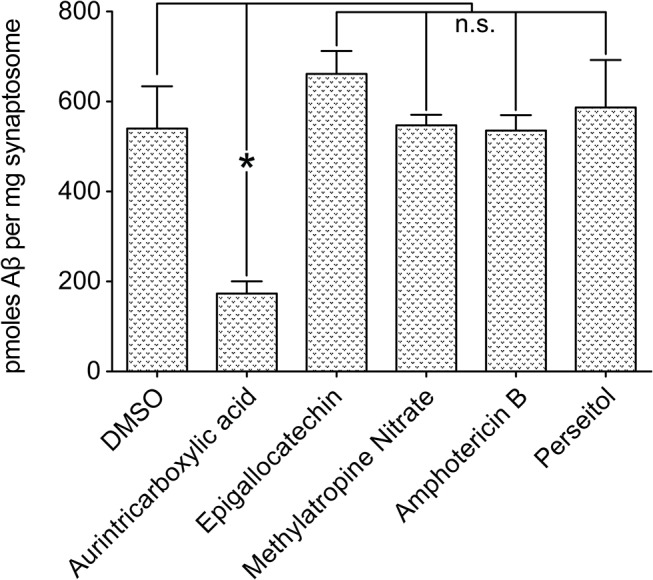
Despite their value as sources of therapeutic drug targets, membrane proteomes are largely inaccessible to high-throughput screening (HTS) tools designed for soluble proteins. An important example comprises the membrane proteins that bind amyloid β oligomers (AβOs). AβOs are neurotoxic ligands thought to instigate the synapse damage that leads to Alzheimer's dementia. At present, the identities of initial AβO binding sites are highly uncertain, largely because of extensive protein-protein interactions that occur following attachment of AβOs to surface membranes. Here, we show that AβO binding sites can be obtained in a state suitable for unbiased HTS by encapsulating the solubilized synaptic membrane proteome into nanoscale lipid bilayers (Nanodiscs). This method gives a soluble membrane protein library (SMPL)--a collection of individualized synaptic proteins in a soluble state. Proteins within SMPL Nanodiscs showed enzymatic and ligand binding activity consistent with conformational integrity. AβOs were found to bind SMPL Nanodiscs with high affinity and specificity, with binding dependent on intact synaptic membrane proteins, and selective for the higher molecular weight oligomers known to accumulate at synapses. Combining SMPL Nanodiscs with a mix-incubate-read chemiluminescence assay provided a solution-based HTS platform to discover antagonists of AβO binding. Screening a library of 2700 drug-like compounds and natural products yielded one compound that potently reduced AβO binding to SMPL Nanodiscs, synaptosomes, and synapses in nerve cell cultures. Although not a therapeutic candidate, this small molecule inhibitor of synaptic AβO binding will provide a useful experimental antagonist for future mechanistic studies of AβOs in Alzheimer's model systems. Overall, results provide proof of concept for using SMPLs in high throughput screening for AβO binding antagonists, and illustrate in general how a SMPL Nanodisc system can facilitate drug discovery for membrane protein targets.
Conflict of interest statement
Figures




References
-
- Cooper MA (2004) Advances in membrane receptor screening and analysis. J Mol Recognit 17: 286–315. - PubMed
-
- Dodgson K, Gedge L, Murray DC, Coldwell M (2009) A 100K well screen for a muscarinic receptor using the Epic label-free system—a reflection on the benefits of the label-free approach to screening seven-transmembrane receptors. J Recept Signal Transduct Res 29: 163–172. 10.1080/10799890903079844 - DOI - PubMed
-
- Gonzalez R, Jennings LL, Knuth M, Orth AP, Klock HE, Ou W, et al. (2010) Screening the mammalian extracellular proteome for regulators of embryonic human stem cell pluripotency. Proceedings of the National Academy of Sciences of the United States of America 107: 3552–3557. 10.1073/pnas.0914019107 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- GM33775/GM/NIGMS NIH HHS/United States
- R21 AG041953/AG/NIA NIH HHS/United States
- R01 AG022547/AG/NIA NIH HHS/United States
- AG029460/AG/NIA NIH HHS/United States
- F32 AG039171/AG/NIA NIH HHS/United States
- R35 GM118145/GM/NIGMS NIH HHS/United States
- AG041953/AG/NIA NIH HHS/United States
- AG020506/AG/NIA NIH HHS/United States
- AG039171/AG/NIA NIH HHS/United States
- AG022547/AG/NIA NIH HHS/United States
- R01 GM033775/GM/NIGMS NIH HHS/United States
- T32 AG020506/AG/NIA NIH HHS/United States
- R01 AG029460/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
