

Activated microglia/macrophage whey acidic protein (AMWAP) inhibits NFκB signaling and induces a neuroprotective phenotype in microglia
- PMID: 25928566
- PMCID: PMC4417279
- DOI: 10.1186/s12974-015-0296-6
Activated microglia/macrophage whey acidic protein (AMWAP) inhibits NFκB signaling and induces a neuroprotective phenotype in microglia
Abstract
Background: Microglia reactivity is a hallmark of neurodegenerative diseases. We have previously identified activated microglia/macrophage whey acidic protein (AMWAP) as a counter-regulator of pro-inflammatory response. Here, we studied its mechanisms of action with a focus on toll-like receptor (TLR) and nuclear factor κB (NFκB) signaling.
Methods: Recombinant AMWAP was produced in Escherichia coli and HEK293 EBNA cells and purified by affinity chromatography. AMWAP uptake was identified by fluorescent labeling, and pro-inflammatory microglia markers were measured by qRT-PCR after stimulation with TLR ligands. NFκB pathway proteins were assessed by immunocytochemistry, Western blot, and immunoprecipitation. A 20S proteasome activity assay was used to investigate the anti-peptidase activity of AMWAP. Microglial neurotoxicity was estimated by nitrite measurement and quantification of caspase 3/7 levels in 661W photoreceptors cultured in the presence of microglia-conditioned medium. Microglial proliferation was investigated using flow cytometry, and their phagocytosis was monitored by the uptake of 661W photoreceptor debris.
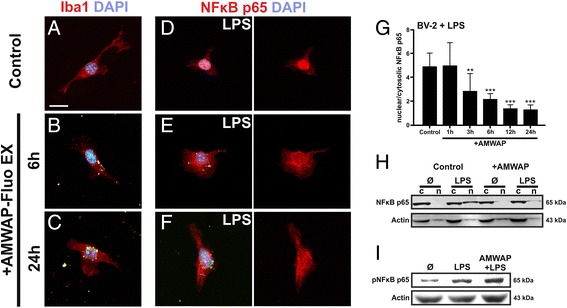
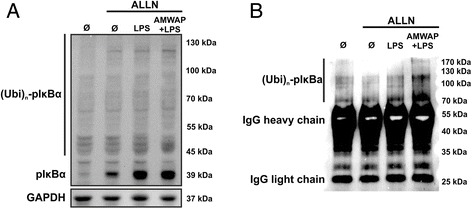
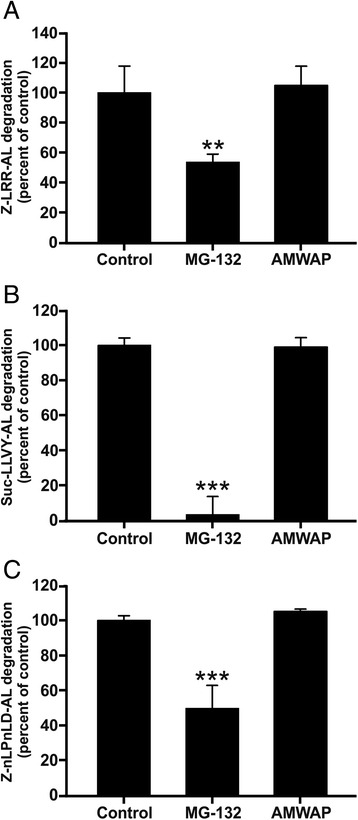
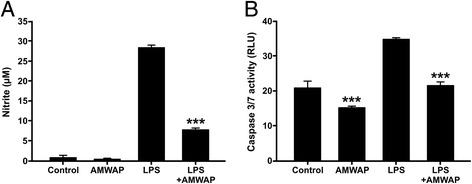
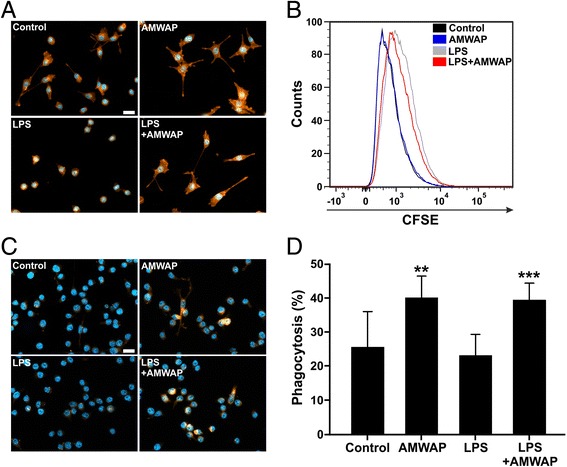
Results: AMWAP was secreted from lipopolysaccharide (LPS)-activated microglia and recombinant AMWAP reduced gene transcription of IL6, iNOS, CCL2, CASP11, and TNFα in BV-2 microglia treated with LPS as TLR4 ligand. This effect was replicated with murine embryonic stem cell-derived microglia (ESdM) and primary brain microglia. AMWAP also diminished pro-inflammatory markers in microglia activated with the TLR2 ligand zymosan but had no effects on IL6, iNOS, and CCL2 transcription in cells treated with CpG oligodeoxynucleotides as TLR9 ligand. Microglial uptake of AMWAP effectively inhibited TLR4-dependent NFκB activation by preventing IRAK-1 and IκBα proteolysis. No inhibition of IκBα phosphorylation or ubiquitination and no influence on overall 20S proteasome activity were observed. Functionally, both microglial nitric oxide (NO) secretion and 661W photoreceptor apoptosis were significantly reduced after AMWAP treatment. AMWAP promoted the filopodia formation of microglia and increased the phagocytic uptake of apoptotic 661W photoreceptor cells.
Conclusions: AMWAP is secreted from reactive microglia and acts in a paracrine fashion to counter-balance TLR2/TLR4-induced reactivity through NFκB inhibition. AMWAP also induces a neuroprotective microglial phenotype with reduced neurotoxicity and increased phagocytosis. We therefore hypothesize that anti-inflammatory whey acidic proteins could have a therapeutic potential in neurodegenerative diseases of the brain and the retina.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
