

Upregulation of cytochrome c oxidase subunit 6b1 (Cox6b1) and formation of mitochondrial supercomplexes: implication of Cox6b1 in the effect of calorie restriction
- PMID: 25929654
- PMCID: PMC4416092
- DOI: 10.1007/s11357-015-9787-8
Upregulation of cytochrome c oxidase subunit 6b1 (Cox6b1) and formation of mitochondrial supercomplexes: implication of Cox6b1 in the effect of calorie restriction
Abstract
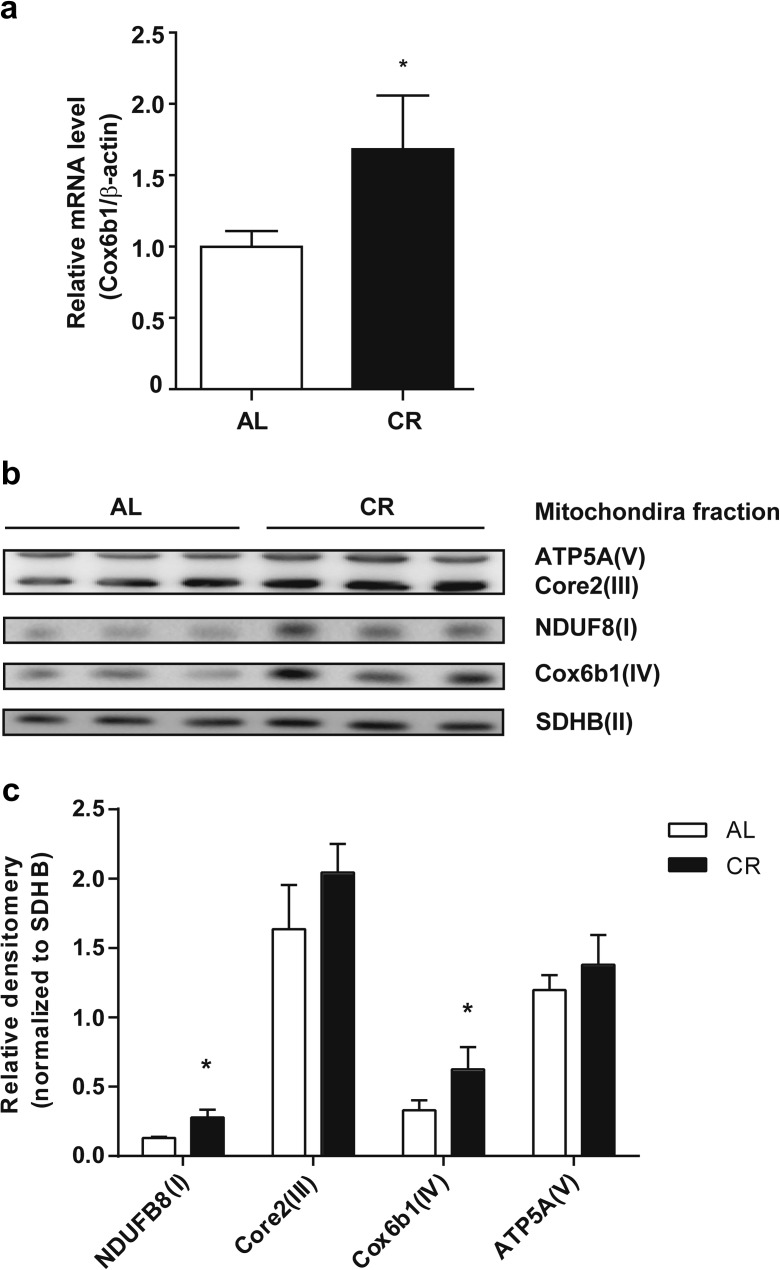
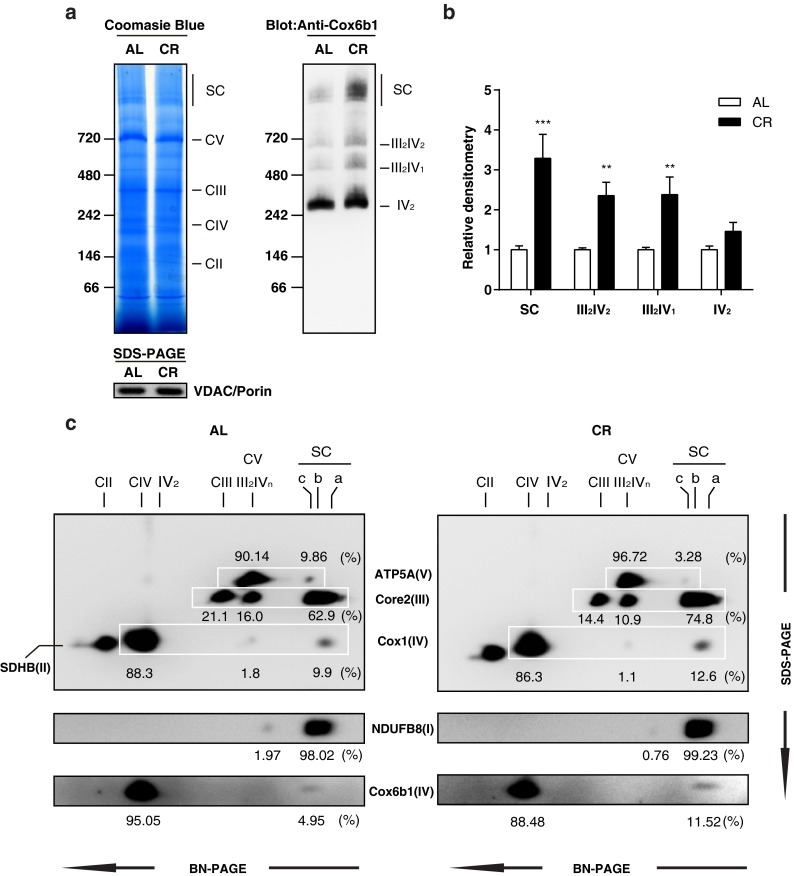
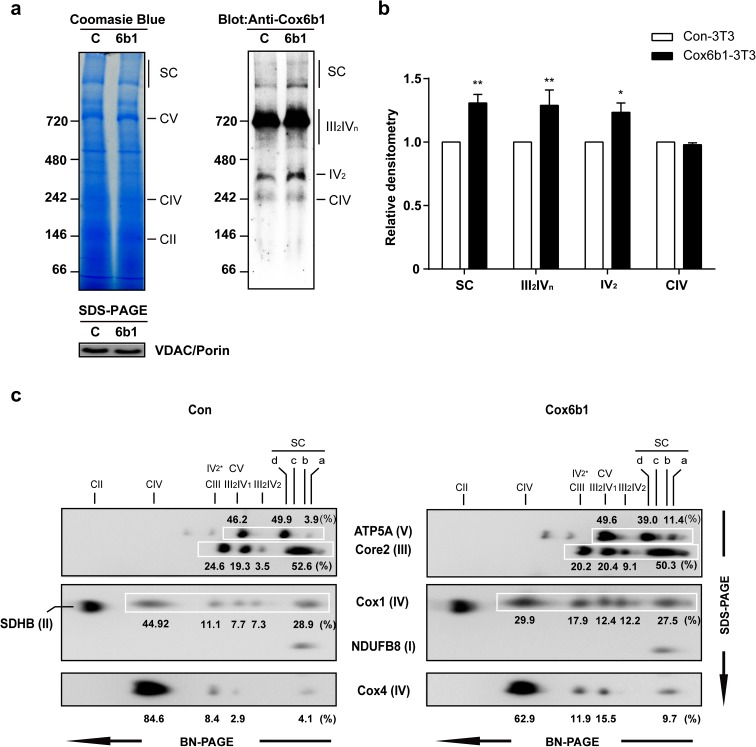
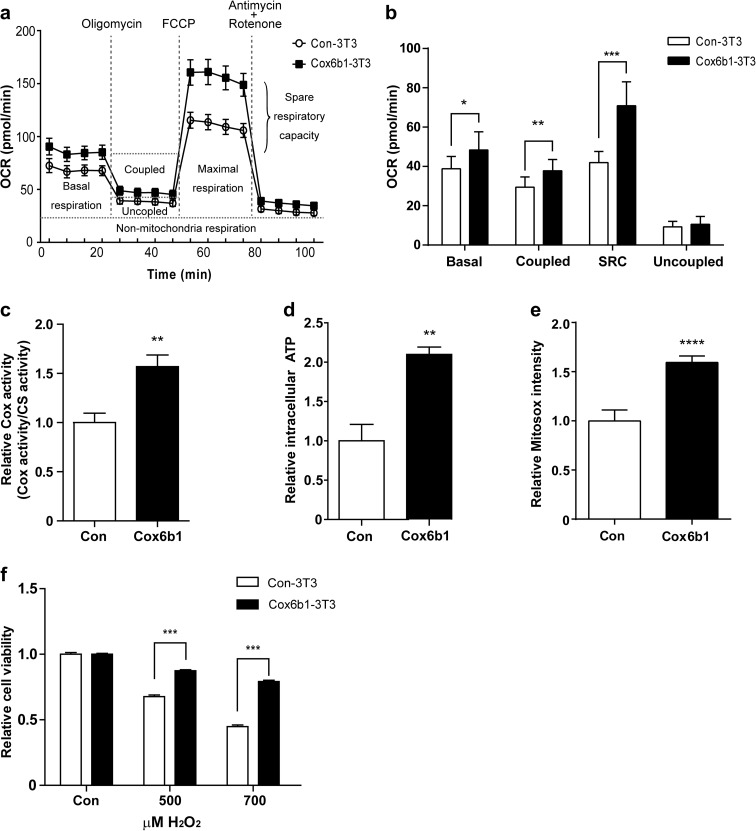
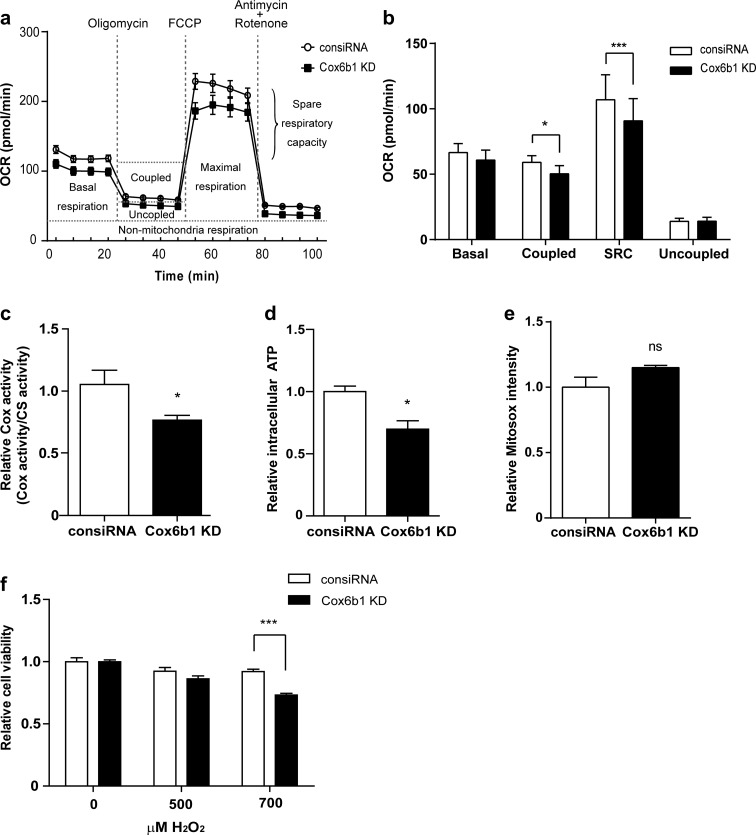
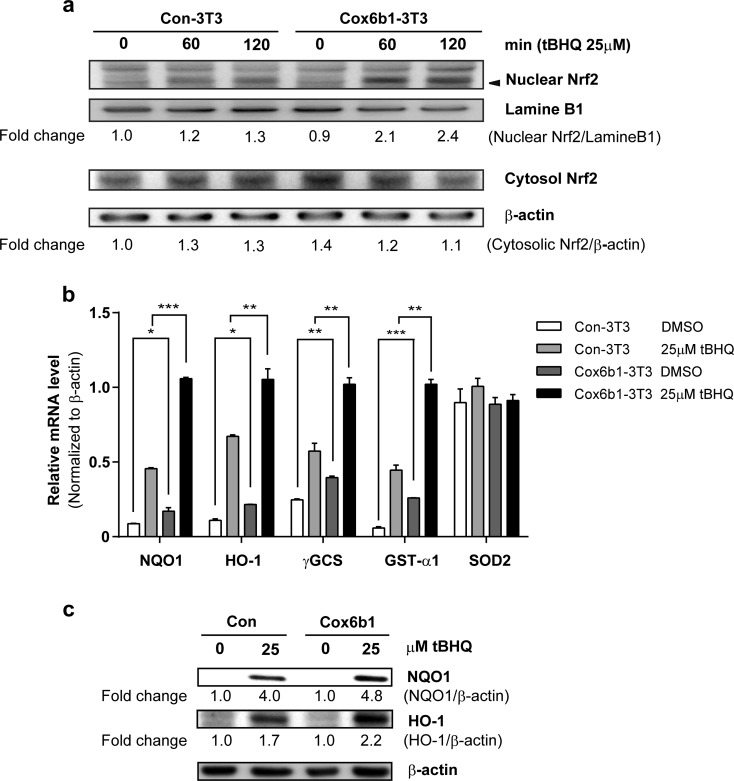
Calorie restriction (CR), a non-genetic intervention that promotes longevity in animals, may exert anti-aging effects by modulating mitochondrial function. Based on our prior mitochondrial proteome analysis, we focused on the potential roles of cytochrome c oxidase (Cox or Complex IV) subunit 6b1 on formation of mitochondrial supercomplexes comprised of Complex I, III, and IV. Blue native polyacrylamide gel electrophoresis followed by immunoblotting showed that the amount of Cox6b1 and the proportion of high molecular weight supercomplexes (SCs) comprised of Complexes I, III, and IV were increased in the liver of mice subjected to 30 % CR, compared with the liver of mice fed ad libitum. In in vitro experiments, in Cox6b1-overexpressing NIH3T3 (Cox6b1-3T3) cells, Cox6b1 was increased in the SC, III2IV1, and III2IV2 complexes and Cox was concomitantly recruited abundantly into the SC, compared with control (Con)-3T3 cells. The proportions of III2IV1, and III2IV2, relative to IV monomer were also increased in Cox6b1-3T3 cells. Cox6b1-3T3 cells showed increased oxygen consumption rates, Cox activity, and intracellular ATP concentrations, indicating enhanced mitochondrial respiration, compared with Con-3T3 cells. Despite the increased basal level of mitochondrial reactive oxygen species (ROS), cell viability after inducing oxidative stress was greater in Cox6b1-3T3 cells than in Con-3T3 cells, probably because of prompt activation of protective mechanisms, such as nuclear translocation of nuclear factor E2-related factor-2. These in vivo and in vitro studies show that Cox6b1 is involved in regulation of mitochondrial function by promoting the formation of SC, suggesting that Cox6b1 contributes to the anti-aging effects of CR.
Figures

Similar articles
-
Role of nuclear-encoded subunit Vb in the assembly and stability of cytochrome c oxidase complex: implications in mitochondrial dysfunction and ROS production.Biochem J. 2009 May 27;420(3):439-49. doi: 10.1042/BJ20090214. Biochem J. 2009. PMID: 19338496 Free PMC article.
-
Mutations in the Yeast Cox12 Subunit Severely Compromise the Activity of the Mitochondrial Complex IV.Biochemistry (Mosc). 2021 Dec;86(12):1607-1623. doi: 10.1134/S0006297921120105. Biochemistry (Mosc). 2021. PMID: 34937540
-
Overexpression of COX6B1 protects against I/R‑induced neuronal injury in rat hippocampal neurons.Mol Med Rep. 2019 Jun;19(6):4852-4862. doi: 10.3892/mmr.2019.10144. Epub 2019 Apr 10. Mol Med Rep. 2019. PMID: 31059068 Free PMC article.
-
The subunit composition and function of mammalian cytochrome c oxidase.Mitochondrion. 2015 Sep;24:64-76. doi: 10.1016/j.mito.2015.07.002. Epub 2015 Jul 17. Mitochondrion. 2015. PMID: 26190566 Review.
-
Regulation of mitochondrial respiration and apoptosis through cell signaling: cytochrome c oxidase and cytochrome c in ischemia/reperfusion injury and inflammation.Biochim Biophys Acta. 2012 Apr;1817(4):598-609. doi: 10.1016/j.bbabio.2011.07.001. Epub 2011 Jul 13. Biochim Biophys Acta. 2012. PMID: 21771582 Free PMC article. Review.
Cited by
-
Specific phytochemicals in floral nectar up-regulate genes involved in longevity regulation and xenobiotic metabolism, extending mosquito life span.Ecol Evol. 2021 May 25;11(12):8363-8380. doi: 10.1002/ece3.7665. eCollection 2021 Jun. Ecol Evol. 2021. PMID: 34188892 Free PMC article.
-
Gut Microbial Postbiotics as Potential Therapeutics for Lymphoma: Proteomics Insights of the Synergistic Effects of Nisin and Urolithin B Against Human Lymphoma Cells.Int J Mol Sci. 2025 Jul 16;26(14):6829. doi: 10.3390/ijms26146829. Int J Mol Sci. 2025. PMID: 40725093 Free PMC article.
-
Astaxanthin Exerts Immunomodulatory Effect by Regulating SDH-HIF-1α Axis and Reprogramming Mitochondrial Metabolism in LPS-Stimulated RAW264.7 Cells.Mar Drugs. 2022 Oct 25;20(11):660. doi: 10.3390/md20110660. Mar Drugs. 2022. PMID: 36354983 Free PMC article.
-
Resilience to capsaicin-induced mitochondrial damage in trigeminal ganglion neurons.Mol Pain. 2020 Jan-Dec;16:1744806920960856. doi: 10.1177/1744806920960856. Mol Pain. 2020. PMID: 32985330 Free PMC article.
-
COX6B1 relieves hypoxia/reoxygenation injury of neonatal rat cardiomyocytes by regulating mitochondrial function.Biotechnol Lett. 2019 Jan;41(1):59-68. doi: 10.1007/s10529-018-2614-4. Epub 2018 Oct 11. Biotechnol Lett. 2019. PMID: 30311029 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources