

Abiotic stress induces change in Cinnamoyl CoA Reductase (CCR) protein abundance and lignin deposition in developing seedlings of Leucaena leucocephala
- PMID: 25931776
- PMCID: PMC4411380
- DOI: 10.1007/s12298-015-0289-z
Abiotic stress induces change in Cinnamoyl CoA Reductase (CCR) protein abundance and lignin deposition in developing seedlings of Leucaena leucocephala
Abstract
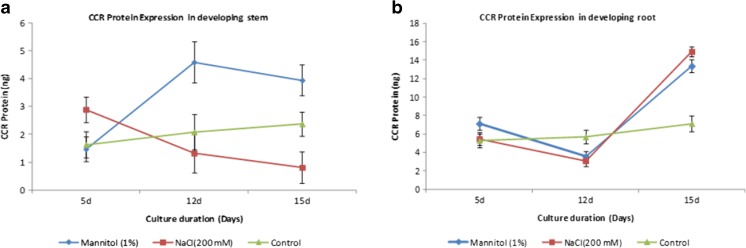
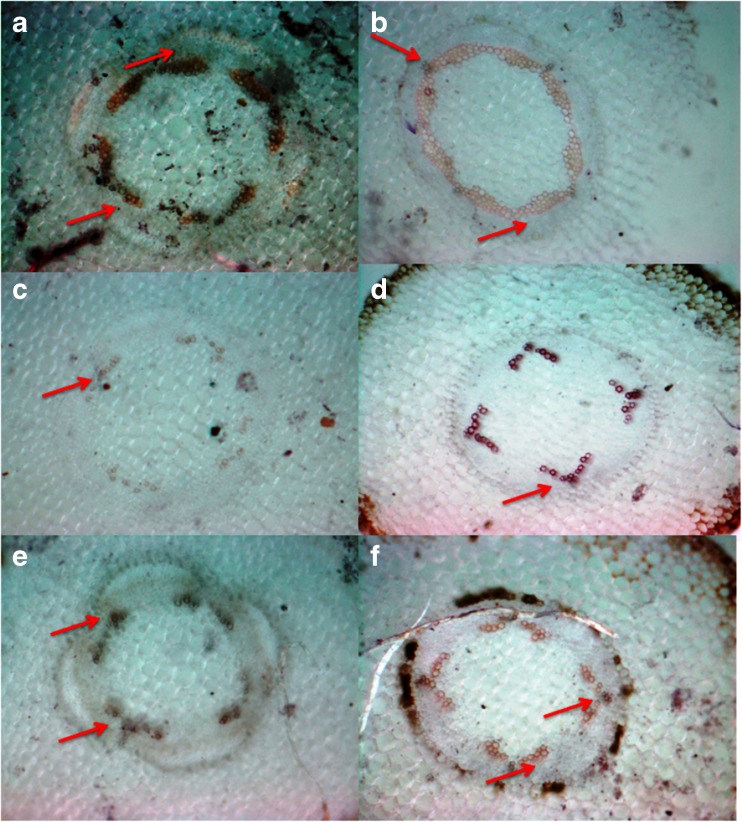
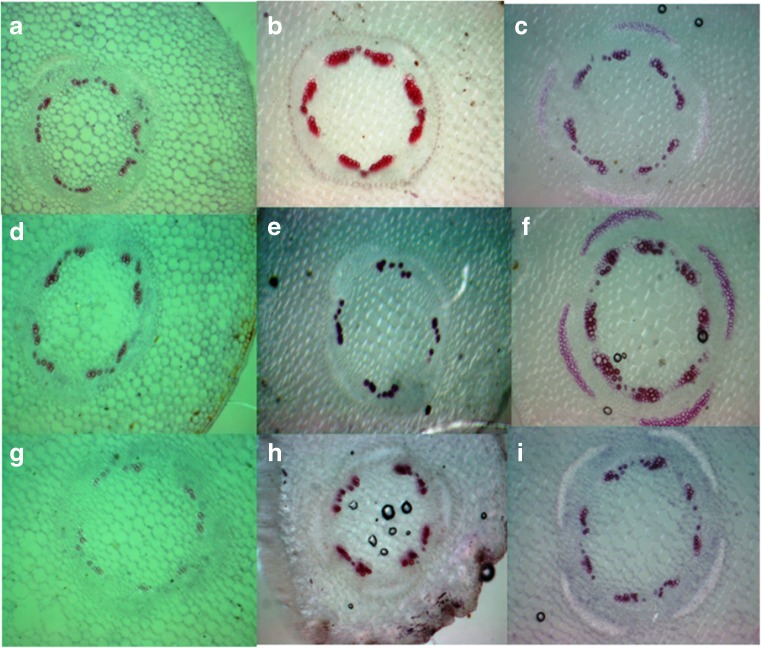
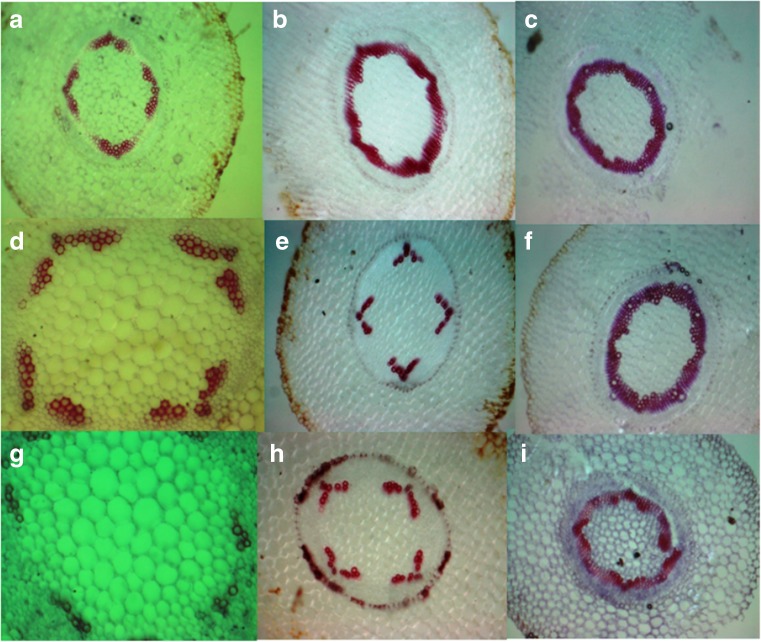
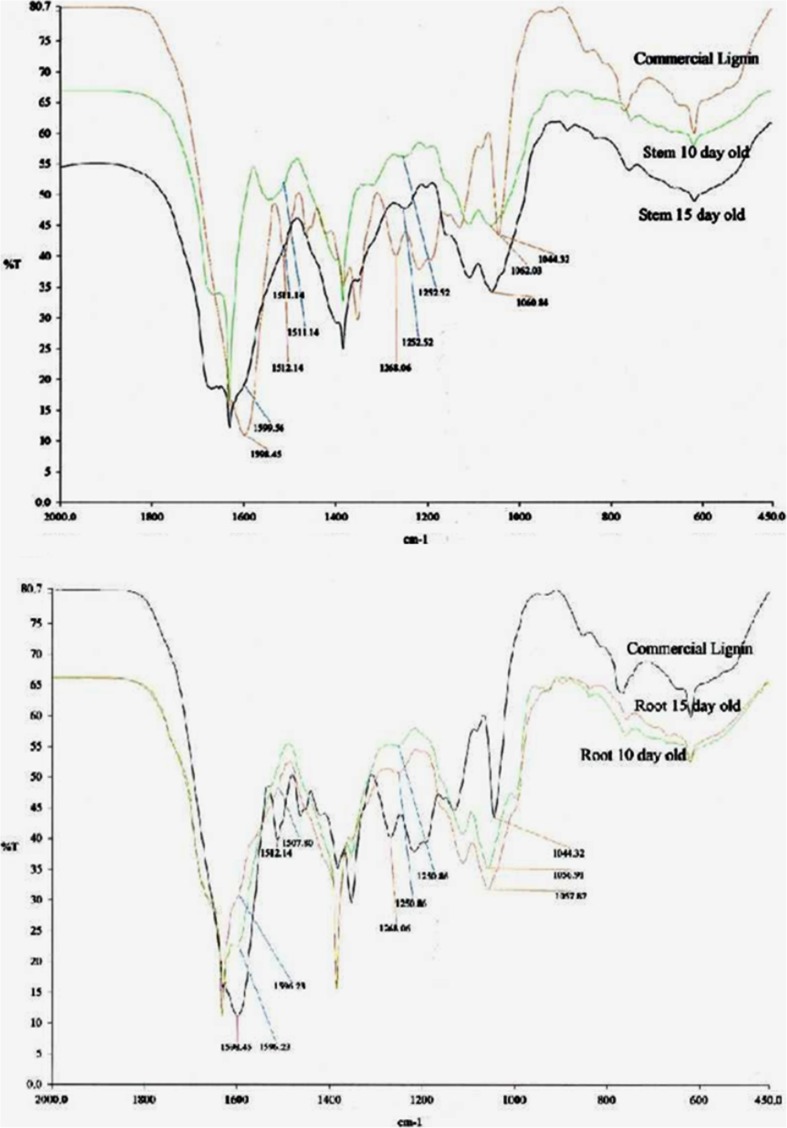
Aboitic stress such as drought and salinity are class of major threats, which plants undergo through their lifetime. Lignin deposition is one of the responses to such abiotic stresses. The gene encoding Cinnamoyl CoA Reductase (CCR) is a key gene for lignin biosynthesis, which has been shown to be over-expressed under stress conditions. In the present study, developing seedlings of Leucaena leucocephala (Vernacular name: Subabul, White popinac) were treated with 1 % mannitol and 200 mM NaCl to mimic drought and salinity stress conditions, respectively. Enzyme linked immunosorbant assay (ELISA) based expression pattern of CCR protein was monitored coupled with Phlorogucinol/HCl activity staining of lignin in transverse sections of developing L. leucocephala seedlings under stress. Our result suggests a differential lignification pattern in developing root and stem under stress conditions. Increase in lignification was observed in mannitol treated stems and corresponding CCR protein accumulation was also higher than control and salt stress treated samples. On the contrary CCR protein was lower in NaCl treated stems and corresponding lignin deposition was also low. Developing root tissue showed a high level of CCR content and lignin deposition than stem samples under all conditions tested. Overall result suggested that lignin accumulation was not affected much in case of developing root however developing stems were significantly affected under drought and salinity stress condition.
Keywords: Abiotic stress; Cinnamoyl CoA reductase; Developing seedlings; Leucaena leucocephala.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources