

Adrenal Development in Mice Requires GATA4 and GATA6 Transcription Factors
- PMID: 25933105
- PMCID: PMC4475720
- DOI: 10.1210/en.2014-1815
Adrenal Development in Mice Requires GATA4 and GATA6 Transcription Factors
Abstract
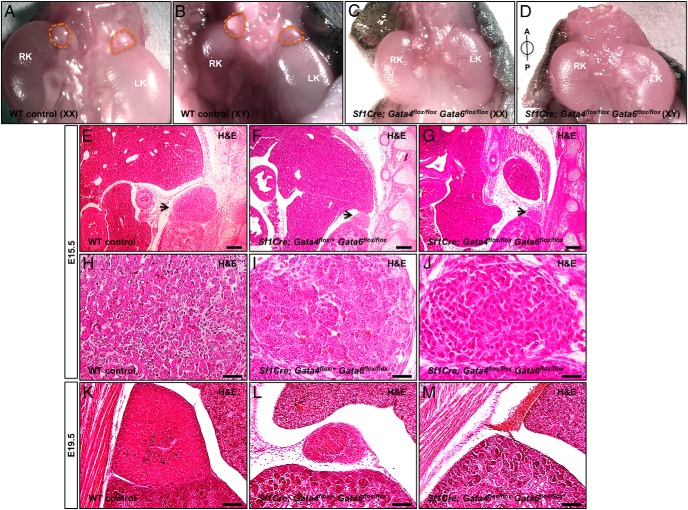
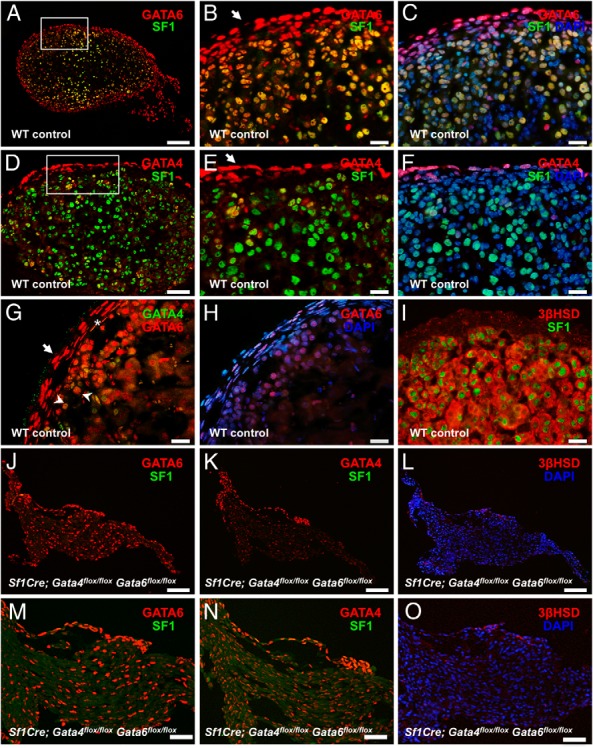
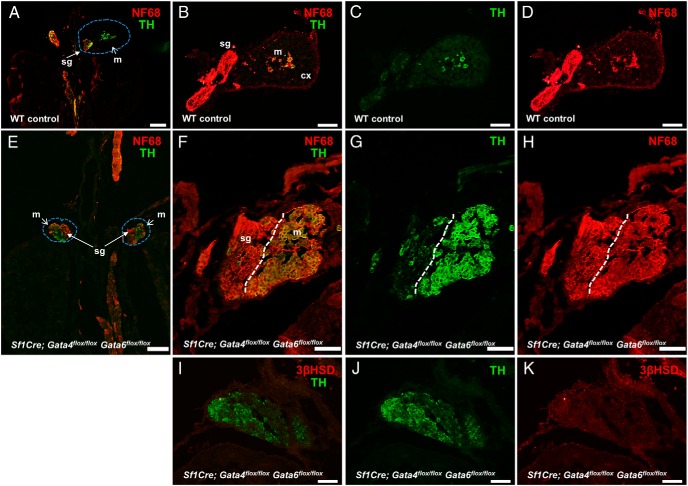
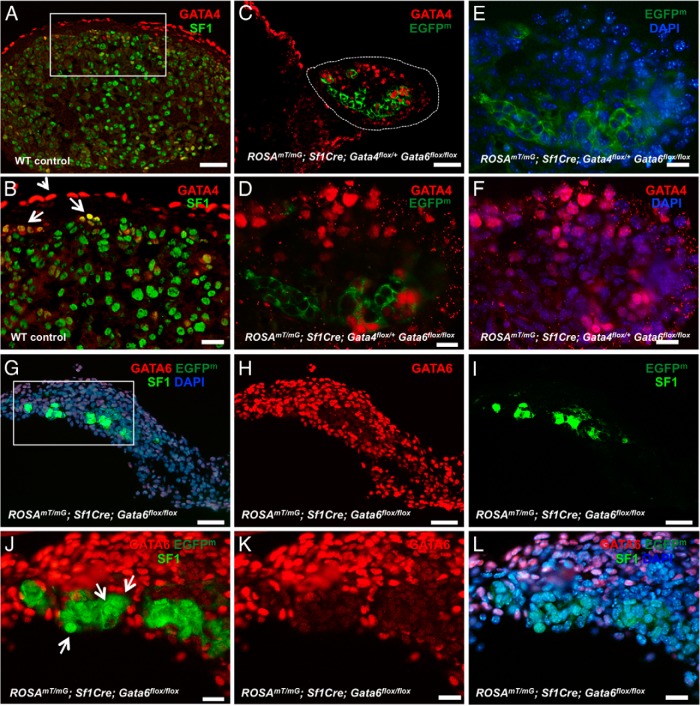
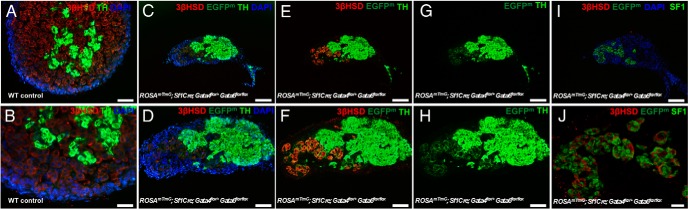
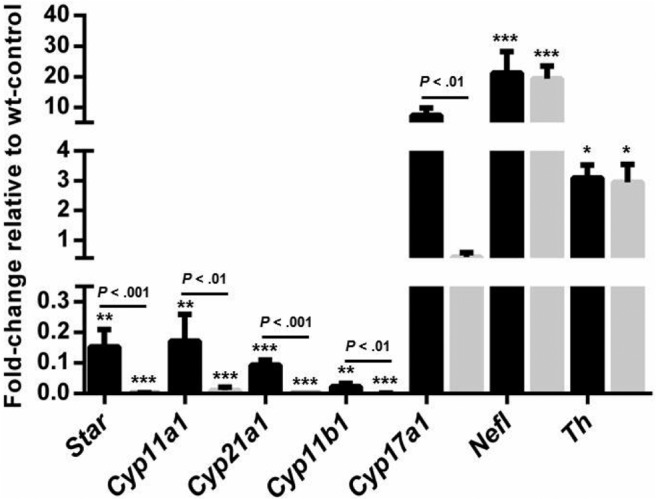
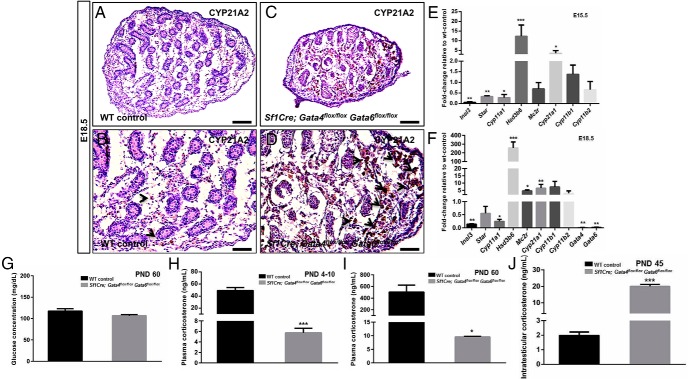
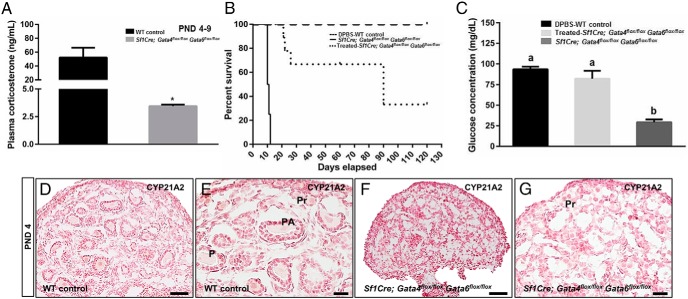
The adrenal glands consist of an outer cortex and an inner medulla, and their primary purposes include hormone synthesis and secretion. The adrenal cortex produces a complex array of steroid hormones, whereas the medulla is part of the sympathetic nervous system and produces the catecholamines epinephrine and norepinephrine. In the mouse, GATA binding protein (GATA) 4 and GATA6 transcription factors are coexpressed in several embryonic tissues, including the adrenal cortex. To explore the roles of GATA4 and GATA6 in mouse adrenal development, we conditionally deleted these genes in adrenocortical cells using the Sf1Cre strain of animals. We report here that mice with Sf1Cre-mediated double deletion of Gata4 and Gata6 genes lack identifiable adrenal glands, steroidogenic factor 1-positive cortical cells and steroidogenic gene expression in the adrenal location. The inactivation of the Gata6 gene alone (Sf1Cre;Gata6(flox/flox)) drastically reduced the adrenal size and corticosterone production in the adult animals. Adrenocortical aplasia is expected to result in the demise of the animal within 2 weeks after birth unless glucocorticoids are provided. In accordance, Sf1Cre;Gata4(flox/flox)Gata6(flox/flox) females depend on steroid supplementation to survive after weaning. Surprisingly, Sf1Cre;Gata4(flox/flox)Gata6(flox/flox) males appear to live normal lifespans as vital steroidogenic synthesis shifts to their testes. Our results reveal a requirement for GATA factors in adrenal development and provide a novel tool to characterize the transcriptional network controlling adrenocortical cell fates.
Figures
Similar articles
-
Combined loss of the GATA4 and GATA6 transcription factors in male mice disrupts testicular development and confers adrenal-like function in the testes.Endocrinology. 2015 May;156(5):1873-86. doi: 10.1210/en.2014-1907. Epub 2015 Feb 10. Endocrinology. 2015. PMID: 25668066 Free PMC article.
-
Testicular steroidogenic cells to the rescue.Endocrinology. 2015 May;156(5):1616-9. doi: 10.1210/en.2015-1222. Endocrinology. 2015. PMID: 25886071 No abstract available.
-
Simultaneous gene deletion of gata4 and gata6 leads to early disruption of follicular development and germ cell loss in the murine ovary.Biol Reprod. 2014 Jul;91(1):24. doi: 10.1095/biolreprod.113.117002. Epub 2014 Jun 4. Biol Reprod. 2014. PMID: 24899573 Free PMC article.
-
Adrenocortical tumorigenesis, luteinizing hormone receptor and transcription factors GATA-4 and GATA-6.Mol Cell Endocrinol. 2007 Apr 15;269(1-2):38-45. doi: 10.1016/j.mce.2006.11.013. Epub 2007 Feb 6. Mol Cell Endocrinol. 2007. PMID: 17337116 Review.
-
Adrenocortical tumorigenesis in transgenic mice: the role of luteinizing hormone receptor and transcription factors GATA-4 and GATA-61.Reprod Biol. 2001 Jul;1(1):5-9. Reprod Biol. 2001. PMID: 14666170 Review.
Cited by
-
Targeted Disruption of Lats1 and Lats2 in Mice Impairs Adrenal Cortex Development and Alters Adrenocortical Cell Fate.Endocrinology. 2020 May 1;161(5):bqaa052. doi: 10.1210/endocr/bqaa052. Endocrinology. 2020. PMID: 32243503 Free PMC article.
-
Transgenic Mouse Models to Study the Development and Maintenance of the Adrenal Cortex.Int J Mol Sci. 2022 Nov 19;23(22):14388. doi: 10.3390/ijms232214388. Int J Mol Sci. 2022. PMID: 36430866 Free PMC article. Review.
-
Functional Zonation of the Adult Mammalian Adrenal Cortex.Front Neurosci. 2016 Jun 15;10:238. doi: 10.3389/fnins.2016.00238. eCollection 2016. Front Neurosci. 2016. PMID: 27378832 Free PMC article. Review.
-
Development of Human Adrenocortical Adenoma (HAA1) Cell Line from Zona Reticularis.Int J Mol Sci. 2022 Dec 29;24(1):584. doi: 10.3390/ijms24010584. Int J Mol Sci. 2022. PMID: 36614027 Free PMC article.
-
The prevalence of CTNNB1 mutations in primary aldosteronism and consequences for clinical outcomes.Sci Rep. 2017 Jan 19;7:39121. doi: 10.1038/srep39121. Sci Rep. 2017. PMID: 28102204 Free PMC article.
References
-
- Muglia L, Jacobson L, Dikkes P, Majzoub JA. Corticotropin-releasing hormone deficiency reveals major fetal but not adult glucocorticoid need. Nature. 1995;373(6513):427–432. - PubMed
-
- Parker KL, Schimmer BP. Steroidogenic factor 1: a key determinant of endocrine development and function. Endocr Rev. 1997;18(3):361–377. - PubMed
-
- Barik J, Marti F, Morel C, et al. Chronic stress triggers social aversion via glucocorticoid receptor in dopaminoceptive neurons. Science. 2013;339(6117):332–335. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
