

Malformations of cortical development and epilepsy
- PMID: 25934463
- PMCID: PMC4448581
- DOI: 10.1101/cshperspect.a022392
Malformations of cortical development and epilepsy
Abstract
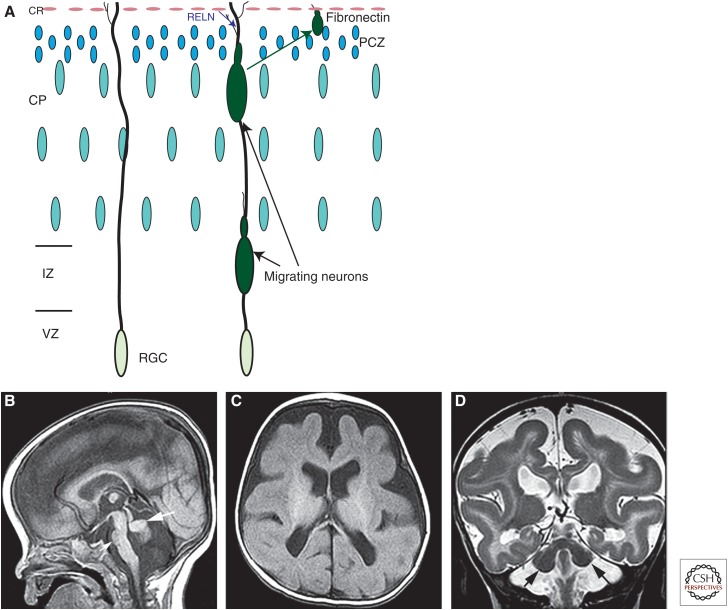
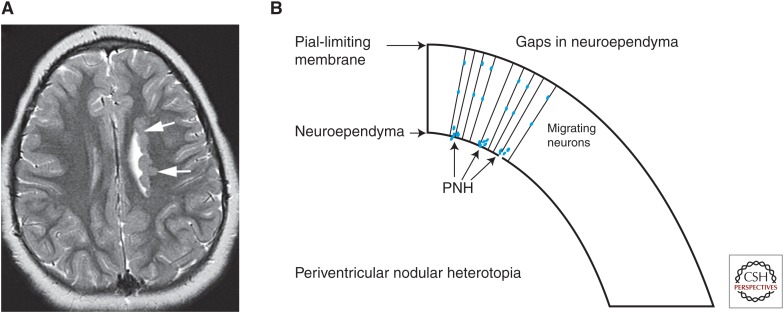
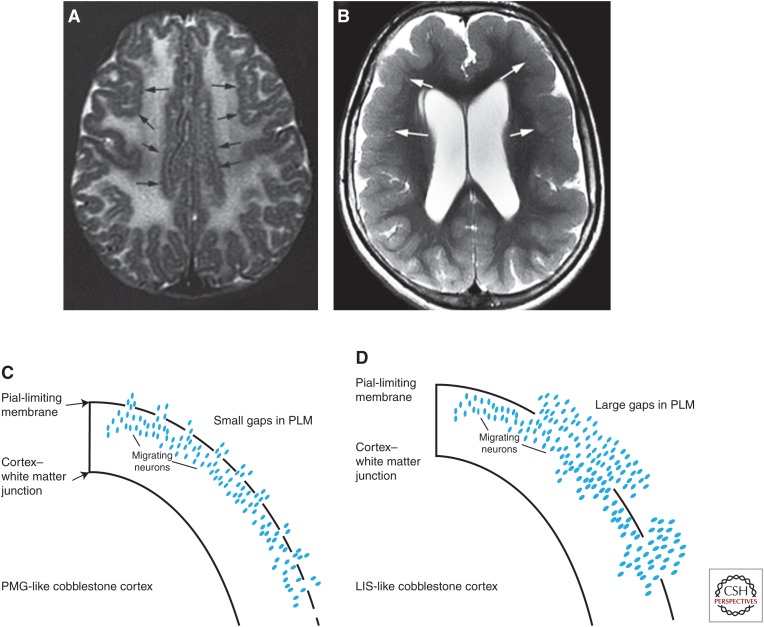
Malformations of cortical development (MCDs) are an important cause of epilepsy and an extremely interesting group of disorders from the perspective of brain development and its perturbations. Many new MCDs have been described in recent years as a result of improvements in imaging, genetic testing, and understanding of the effects of mutations on the ability of their protein products to correctly function within the molecular pathways by which the brain functions. In this review, most of the major MCDs are reviewed from a clinical, embryological, and genetic perspective. The most recent literature regarding clinical diagnosis, mechanisms of development, and future paths of research are discussed.
Copyright © 2015 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
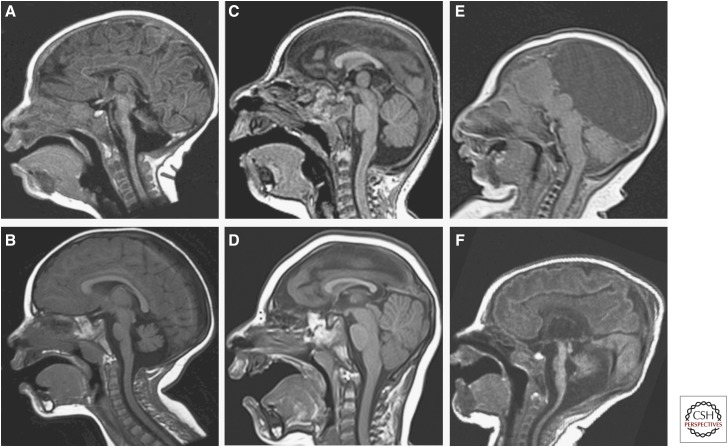
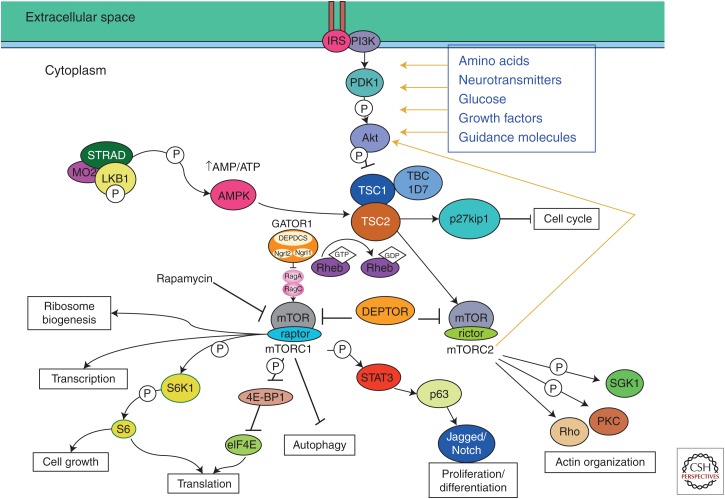
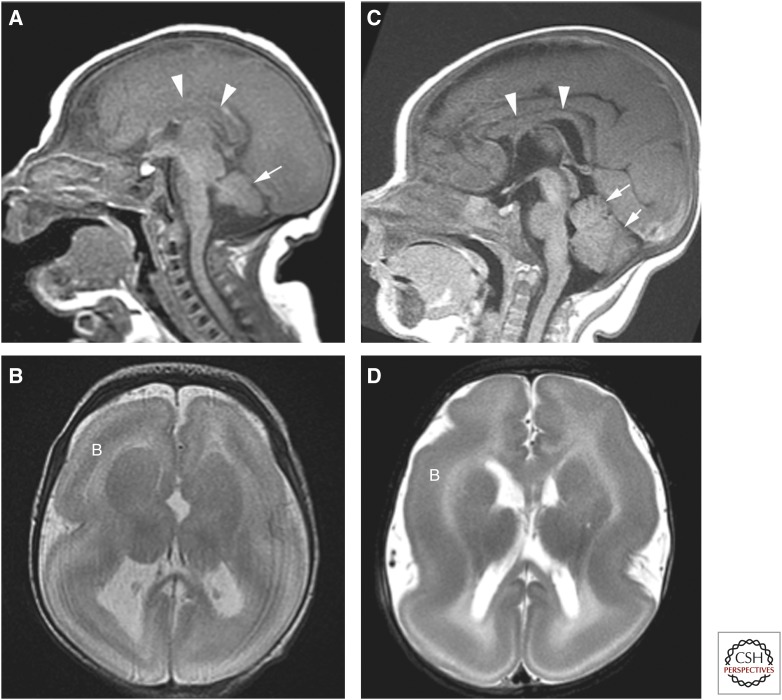
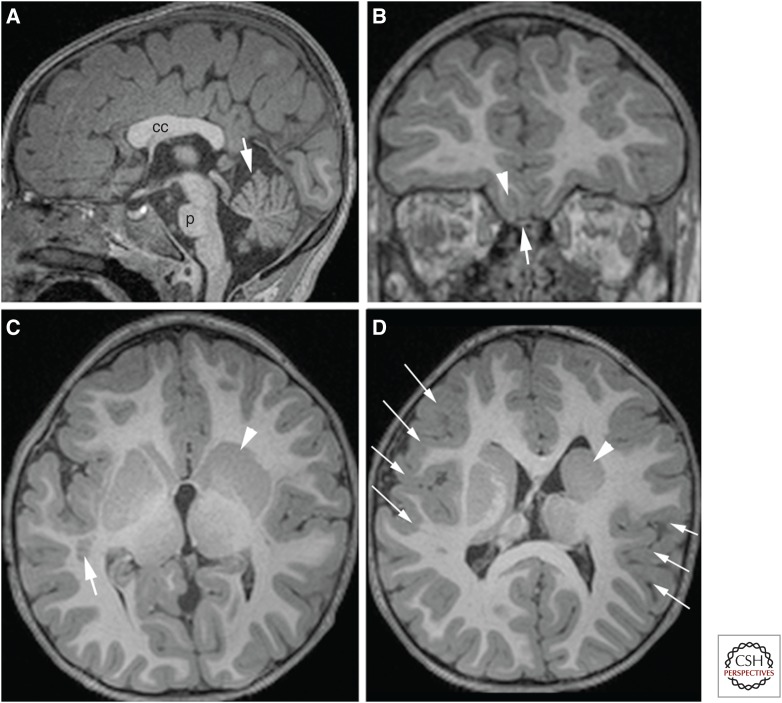
References
-
- Al-Dosari MS, Shaheen R, Colak D, Alkuraya FS. 2010. Novel CENPJ mutation causes Seckel Syndrome. J Med Genet 47: 411–414. - PubMed
-
- Bahi-Buisson N, Poirier K, Boddaert N, Saillour Y, Castelnau L, Philip N, Buyse G, Villard L, Joriot S, Marret S, et al. 2008. Refinement of cortical dysgeneses spectrum associated with TUBA1A mutations. J Med Genet 45: 647–653. - PubMed
-
- Bahi-Buisson N, Poirier K, Boddaert N, Fallet-Bianco C, Specchio N, Bertini E, Caglayan O, Lascelles K, Elie C, Rambaud J, et al. 2010. Gpr56-related bilateral frontoparietal polymicrogyria: Further evidence for an overlap with the cobblestone complex. Brain 133: 3194–3209. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical