

A New DREADD Facilitates the Multiplexed Chemogenetic Interrogation of Behavior
- PMID: 25937170
- PMCID: PMC4441592
- DOI: 10.1016/j.neuron.2015.03.065
A New DREADD Facilitates the Multiplexed Chemogenetic Interrogation of Behavior
Abstract
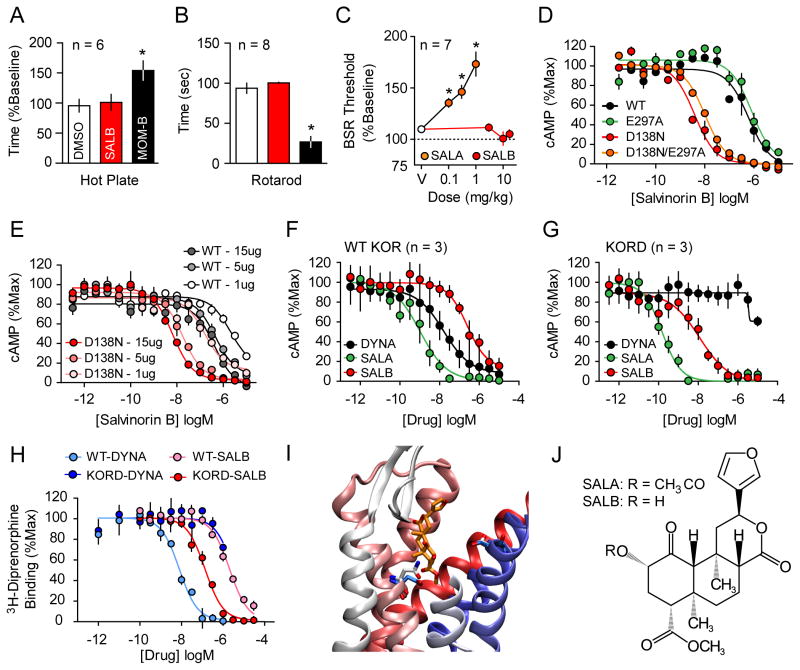
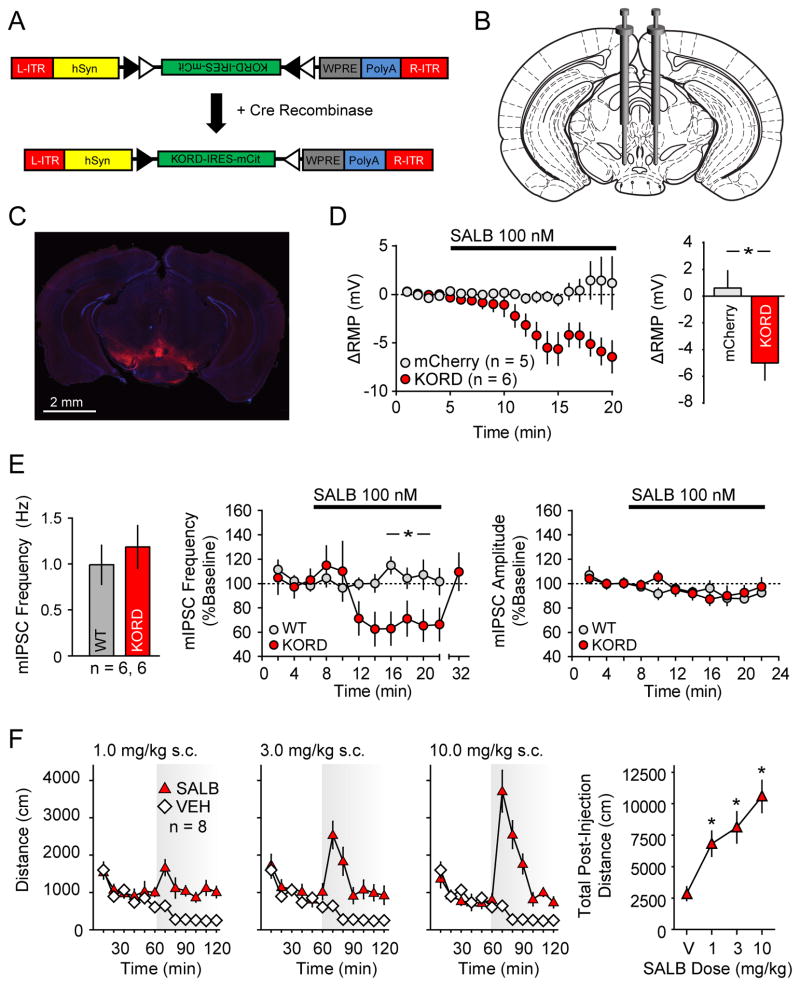
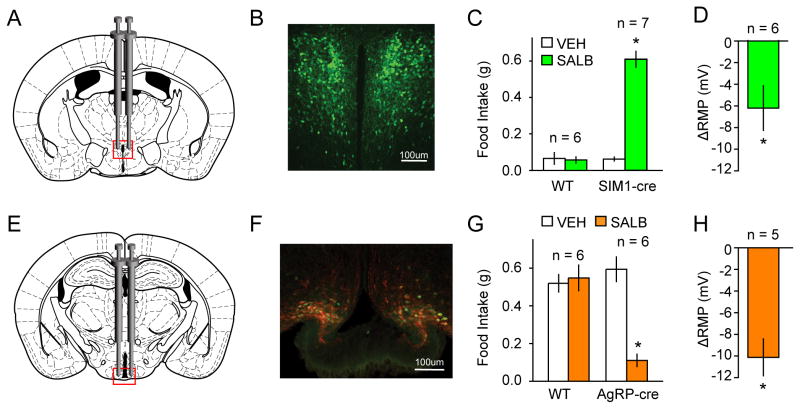
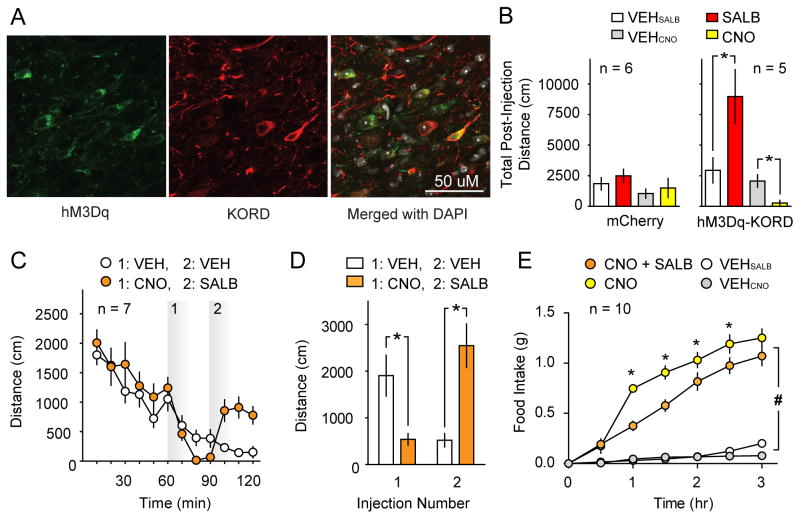
DREADDs are chemogenetic tools widely used to remotely control cellular signaling, neuronal activity, and behavior. Here we used a structure-based approach to develop a new Gi-coupled DREADD using the kappa-opioid receptor as a template (KORD) that is activated by the pharmacologically inert ligand salvinorin B (SALB). Activation of virally expressed KORD in several neuronal contexts robustly attenuated neuronal activity and modified behaviors. Additionally, co-expression of the KORD and the Gq-coupled M3-DREADD within the same neuronal population facilitated the sequential and bidirectional remote control of behavior. The availability of DREADDs activated by different ligands provides enhanced opportunities for investigating diverse physiological systems using multiplexed chemogenetic actuators.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Using opioid receptors to expand the chemogenetic and optogenetic toolbox.Neuron. 2015 May 20;86(4):853-855. doi: 10.1016/j.neuron.2015.05.014. Neuron. 2015. PMID: 25996128
-
Chemogenetic manipulation of neurons.Nat Methods. 2015 Jul;12(7):603. doi: 10.1038/nmeth.3471. Nat Methods. 2015. PMID: 26339707 No abstract available.
References
-
- Ansonoff MA, Zhang J, Czyzyk T, Rothman RB, Stewart J, Xu H, Zjwiony J, Siebert DJ, Yang F, Roth BL, Pintar JE. Antinociceptive and hypothermic effects of Salvinorin A are abolished in a novel strain of kappa-opioid receptor-1 knockout mice. J Pharmacol Exp Ther. 2006;318:641–648. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 MH105892/MH/NIMH NIH HHS/United States
- U01 AA020911/AA/NIAAA NIH HHS/United States
- R01AA019454/AA/NIAAA NIH HHS/United States
- P60 AA011605/AA/NIAAA NIH HHS/United States
- F30 AA021312/AA/NIAAA NIH HHS/United States
- R00 AA017668/AA/NIAAA NIH HHS/United States
- U01AA020911/AA/NIAAA NIH HHS/United States
- R01 DA017204/DA/NIDA NIH HHS/United States
- T32 GM007040/GM/NIGMS NIH HHS/United States
- DK075087/DK/NIDDK NIH HHS/United States
- P30 NS045892/NS/NINDS NIH HHS/United States
- UO1MH105892/MH/NIMH NIH HHS/United States
- F31AA02228001/AA/NIAAA NIH HHS/United States
- Intramural NIH HHS/United States
- DK075089/DK/NIDDK NIH HHS/United States
- P01DA035764/DA/NIDA NIH HHS/United States
- P01 DA035764/DA/NIDA NIH HHS/United States
- R00AA017668/AA/NIAAA NIH HHS/United States
- R01 AA018335/AA/NIAAA NIH HHS/United States
- R01 AA019454/AA/NIAAA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
