

A Potential Contributory Role for Ciliary Dysfunction in the 16p11.2 600 kb BP4-BP5 Pathology
- PMID: 25937446
- PMCID: PMC4570289
- DOI: 10.1016/j.ajhg.2015.04.002
A Potential Contributory Role for Ciliary Dysfunction in the 16p11.2 600 kb BP4-BP5 Pathology
Abstract
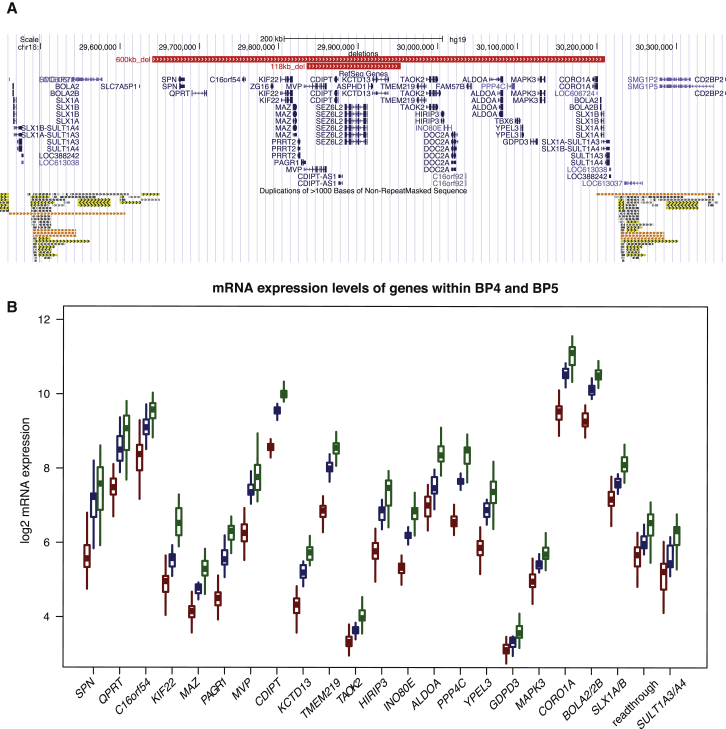
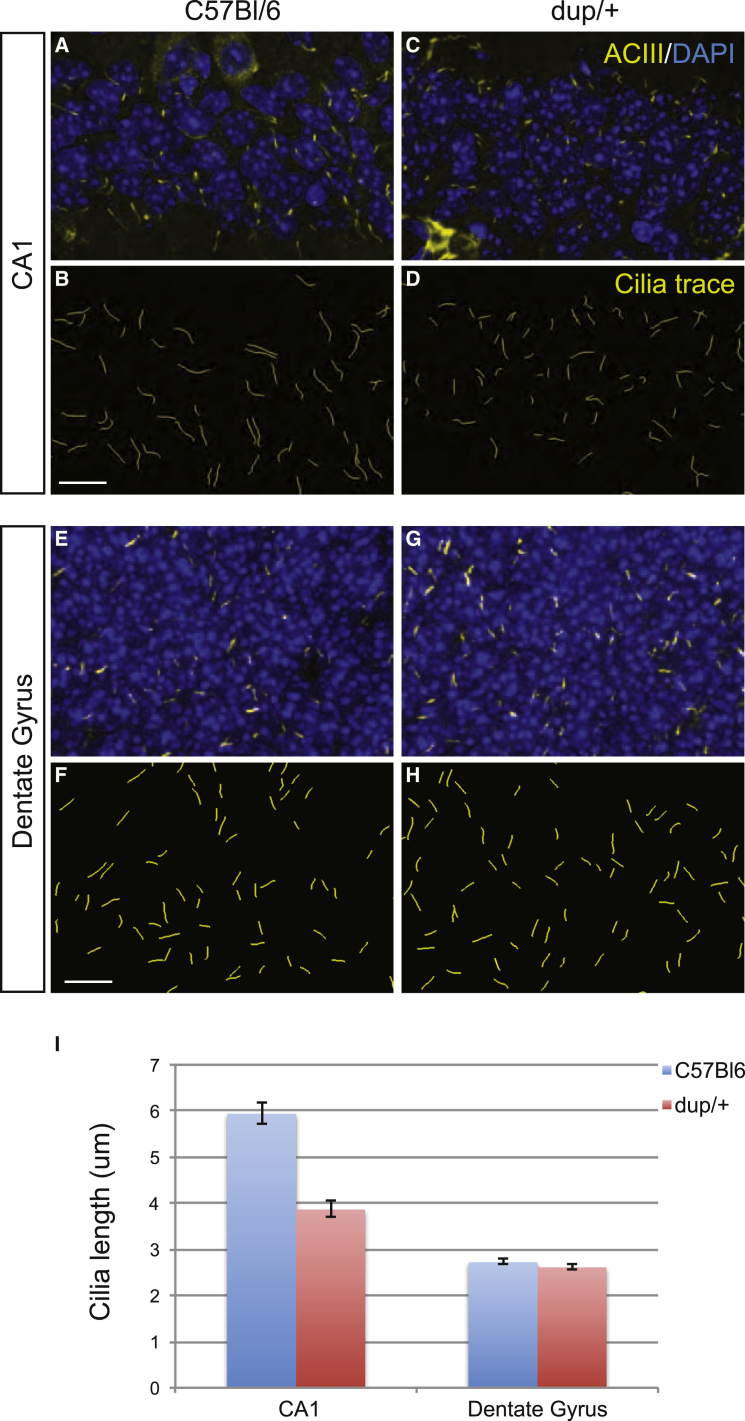
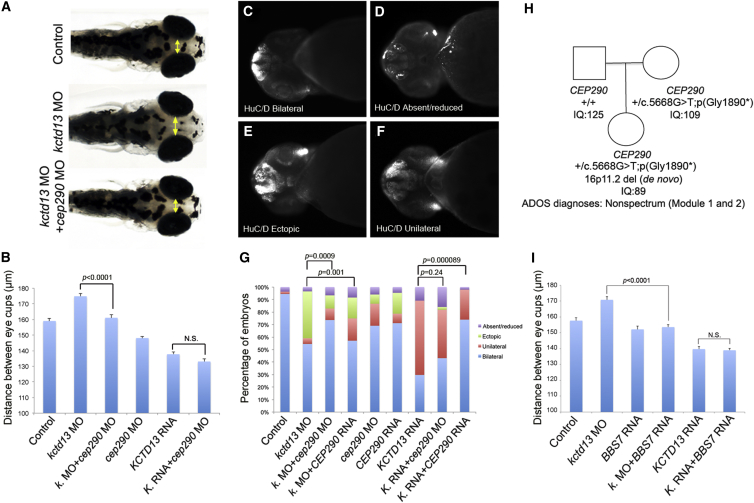
The 16p11.2 600 kb copy-number variants (CNVs) are associated with mirror phenotypes on BMI, head circumference, and brain volume and represent frequent genetic lesions in autism spectrum disorders (ASDs) and schizophrenia. Here we interrogated the transcriptome of individuals carrying reciprocal 16p11.2 CNVs. Transcript perturbations correlated with clinical endophenotypes and were enriched for genes associated with ASDs, abnormalities of head size, and ciliopathies. Ciliary gene expression was also perturbed in orthologous mouse models, raising the possibility that ciliary dysfunction contributes to 16p11.2 pathologies. In support of this hypothesis, we found structural ciliary defects in the CA1 hippocampal region of 16p11.2 duplication mice. Moreover, by using an established zebrafish model, we show genetic interaction between KCTD13, a key driver of the mirrored neuroanatomical phenotypes of the 16p11.2 CNV, and ciliopathy-associated genes. Overexpression of BBS7 rescues head size and neuroanatomical defects of kctd13 morphants, whereas suppression or overexpression of CEP290 rescues phenotypes induced by KCTD13 under- or overexpression, respectively. Our data suggest that dysregulation of ciliopathy genes contributes to the clinical phenotypes of these CNVs.
Copyright © 2015 The American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Weiss L.A., Shen Y., Korn J.M., Arking D.E., Miller D.T., Fossdal R., Saemundsen E., Stefansson H., Ferreira M.A., Green T., Autism Consortium Association between microdeletion and microduplication at 16p11.2 and autism. N. Engl. J. Med. 2008;358:667–675. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
