

Histone H3.3 is required for endogenous retroviral element silencing in embryonic stem cells
- PMID: 25938714
- PMCID: PMC4509593
- DOI: 10.1038/nature14345
Histone H3.3 is required for endogenous retroviral element silencing in embryonic stem cells
Abstract
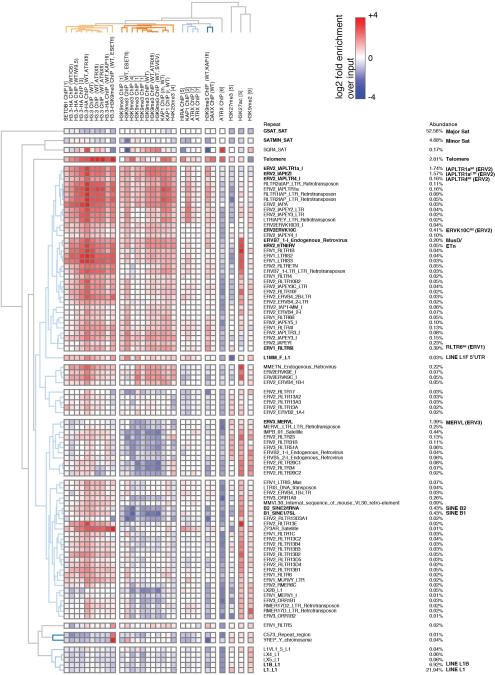
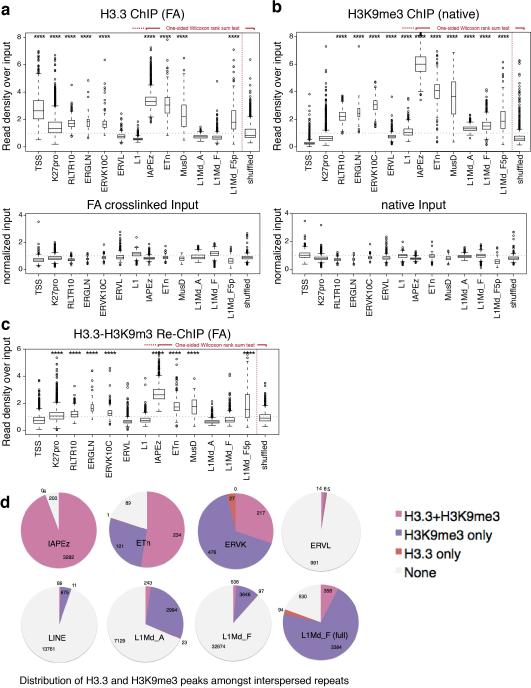
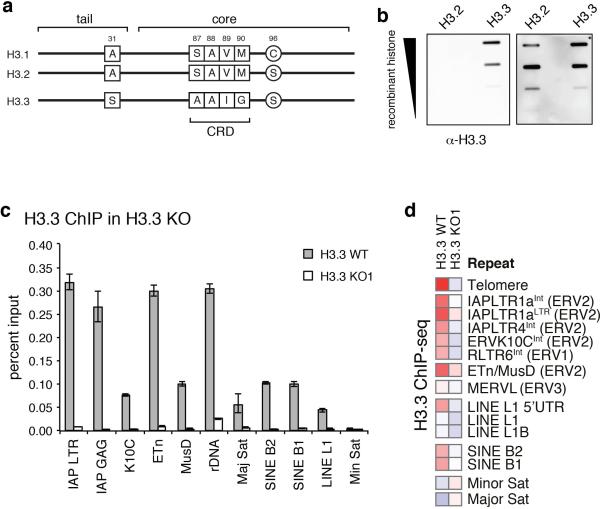
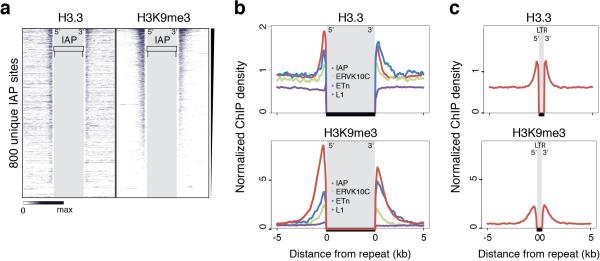
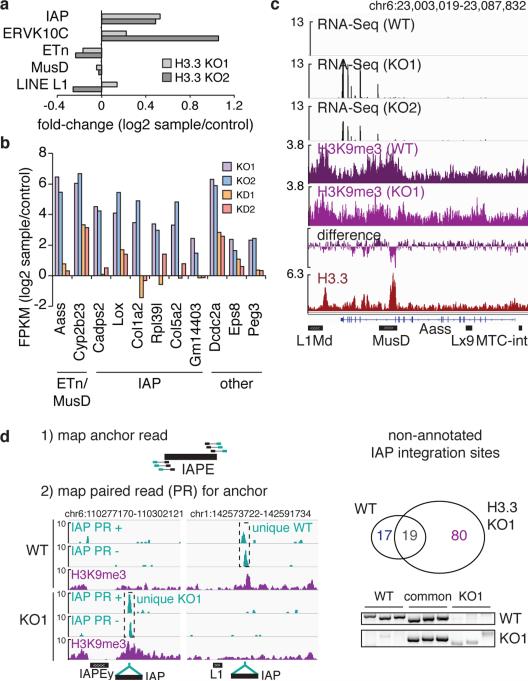
Transposable elements comprise roughly 40% of mammalian genomes. They have an active role in genetic variation, adaptation and evolution through the duplication or deletion of genes or their regulatory elements, and transposable elements themselves can act as alternative promoters for nearby genes, resulting in non-canonical regulation of transcription. However, transposable element activity can lead to detrimental genome instability, and hosts have evolved mechanisms to silence transposable element mobility appropriately. Recent studies have demonstrated that a subset of transposable elements, endogenous retroviral elements (ERVs) containing long terminal repeats (LTRs), are silenced through trimethylation of histone H3 on lysine 9 (H3K9me3) by ESET (also known as SETDB1 or KMT1E) and a co-repressor complex containing KRAB-associated protein 1 (KAP1; also known as TRIM28) in mouse embryonic stem cells. Here we show that the replacement histone variant H3.3 is enriched at class I and class II ERVs, notably those of the early transposon (ETn)/MusD family and intracisternal A-type particles (IAPs). Deposition at a subset of these elements is dependent upon the H3.3 chaperone complex containing α-thalassaemia/mental retardation syndrome X-linked (ATRX) and death-domain-associated protein (DAXX). We demonstrate that recruitment of DAXX, H3.3 and KAP1 to ERVs is co-dependent and occurs upstream of ESET, linking H3.3 to ERV-associated H3K9me3. Importantly, H3K9me3 is reduced at ERVs upon H3.3 deletion, resulting in derepression and dysregulation of adjacent, endogenous genes, along with increased retrotransposition of IAPs. Our study identifies a unique heterochromatin state marked by the presence of both H3.3 and H3K9me3, and establishes an important role for H3.3 in control of ERV retrotransposition in embryonic stem cells.
Figures
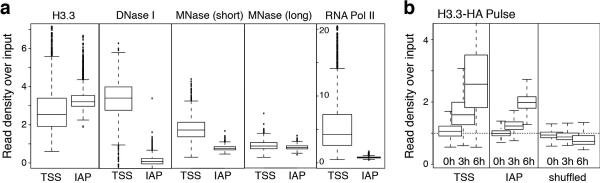

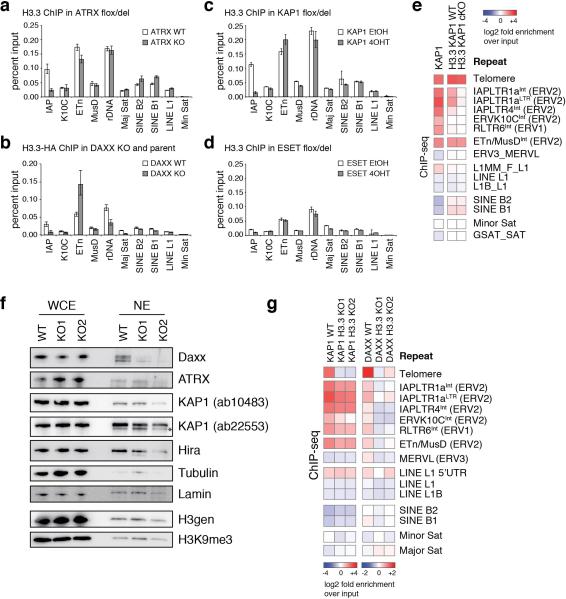
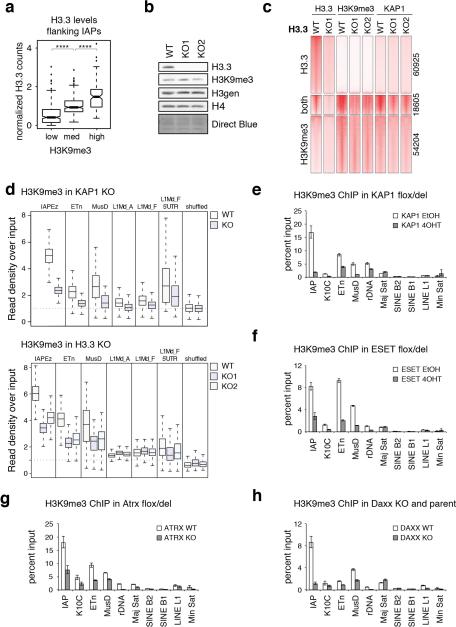
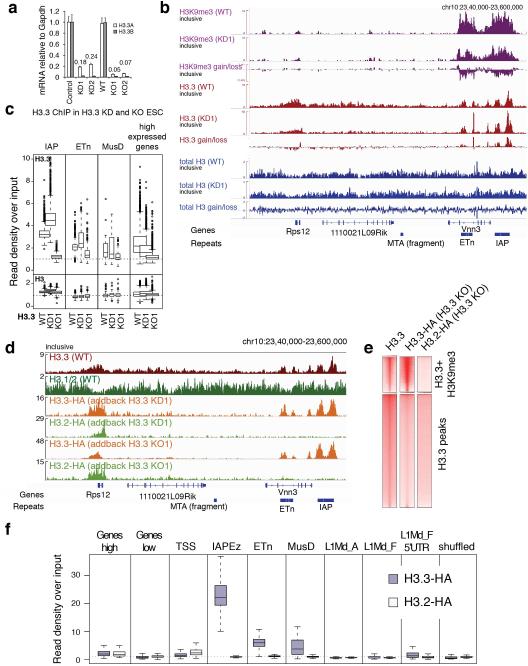
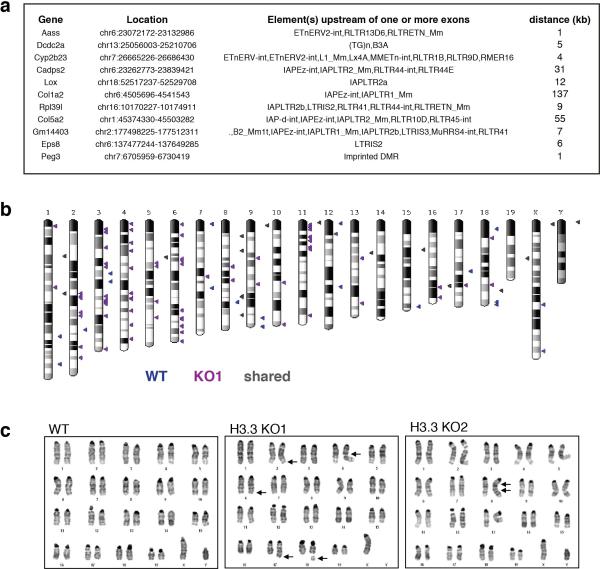
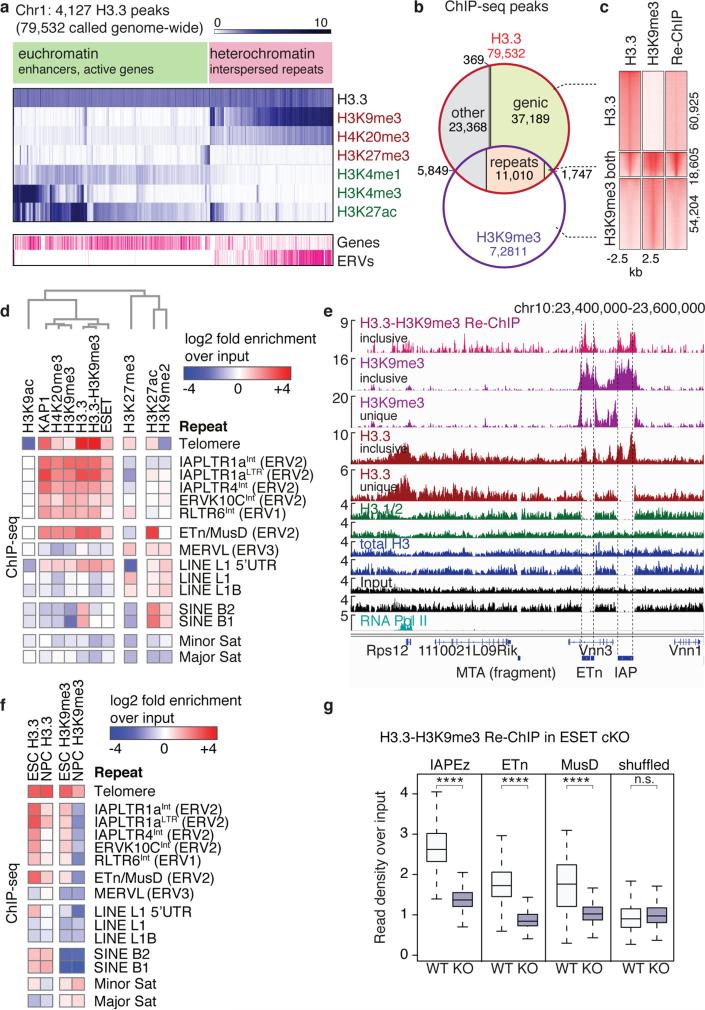
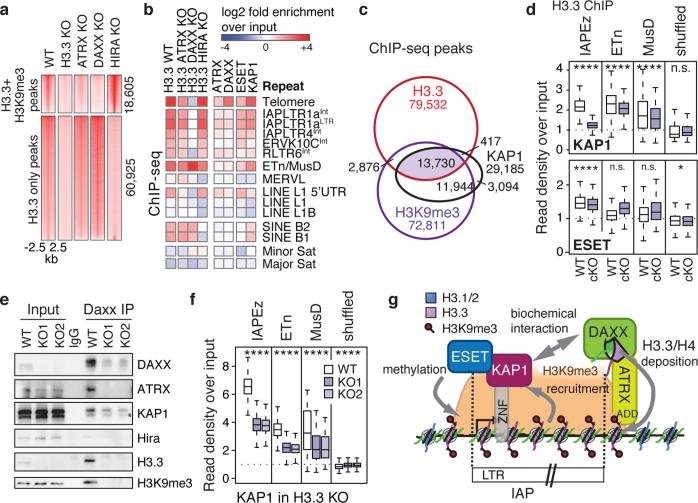
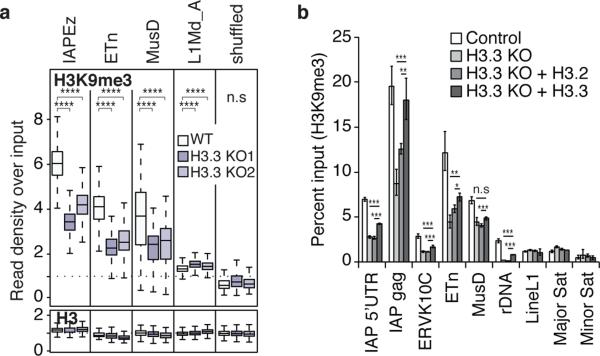
Comment in
-
On the role of H3.3 in retroviral silencing.Nature. 2017 Aug 2;548(7665):E1-E3. doi: 10.1038/nature23277. Nature. 2017. PMID: 28770848 Free PMC article. No abstract available.
References
-
- Waterston RH, et al. Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420:520–562. doi:10.1038/nature01262. - PubMed
-
- Feschotte C, Gilbert C. Endogenous viruses: insights into viral evolution and impact on host biology. Nat Rev Genet. 2012;13:283–296. doi:10.1038/nrg3199. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
