

Obstruction of adaptation in diploids by recessive, strongly deleterious alleles
- PMID: 25941393
- PMCID: PMC4443376
- DOI: 10.1073/pnas.1424949112
Obstruction of adaptation in diploids by recessive, strongly deleterious alleles
Abstract
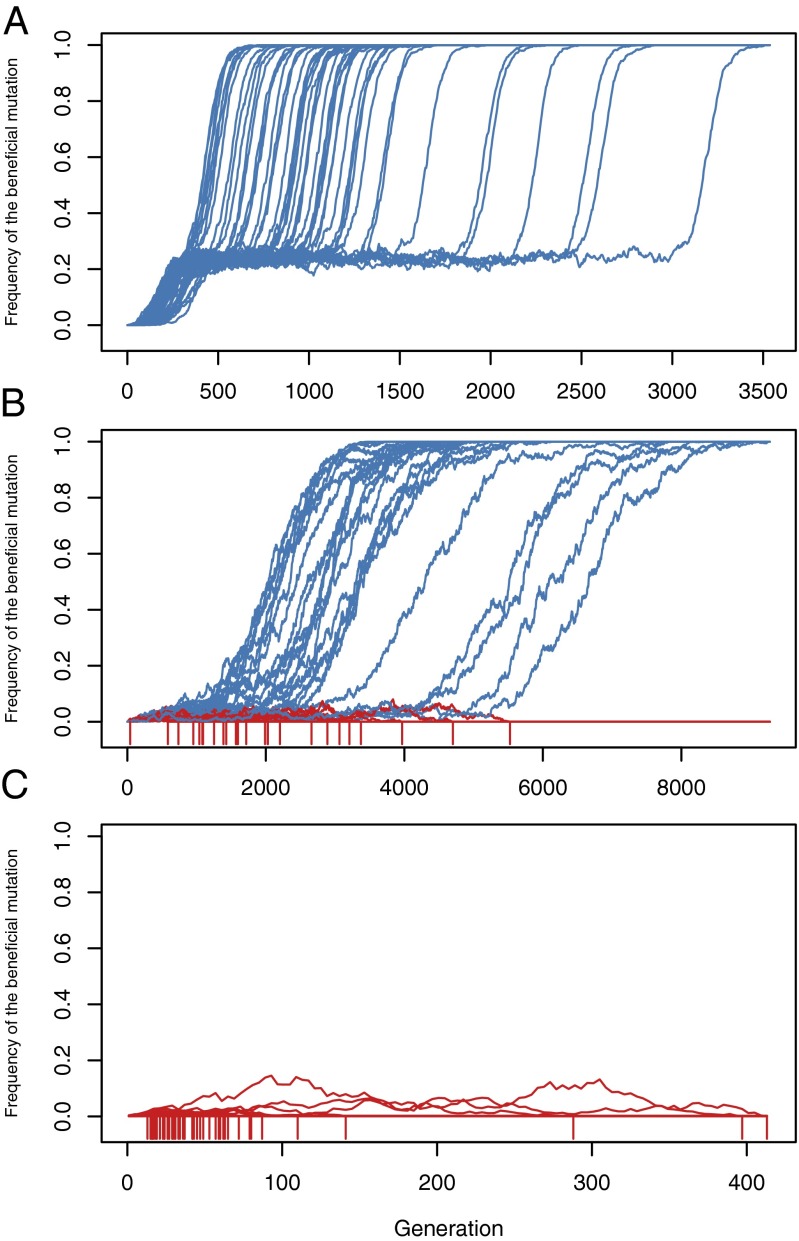
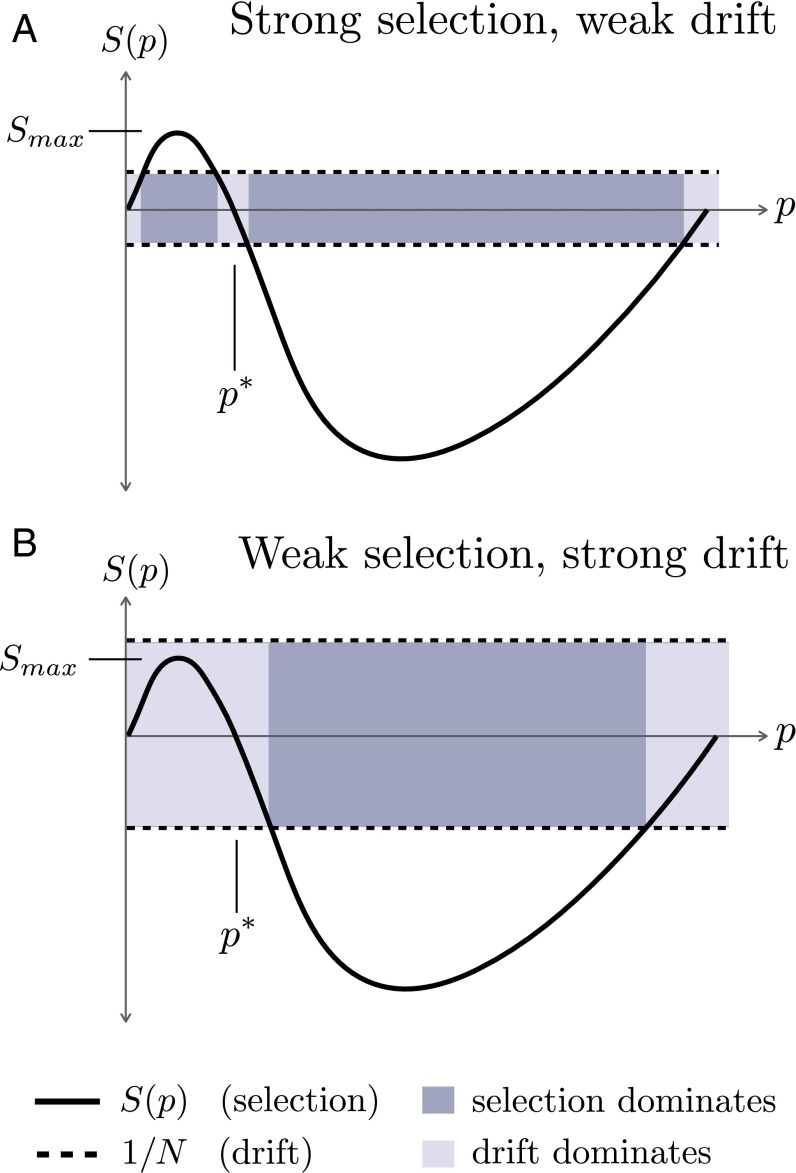
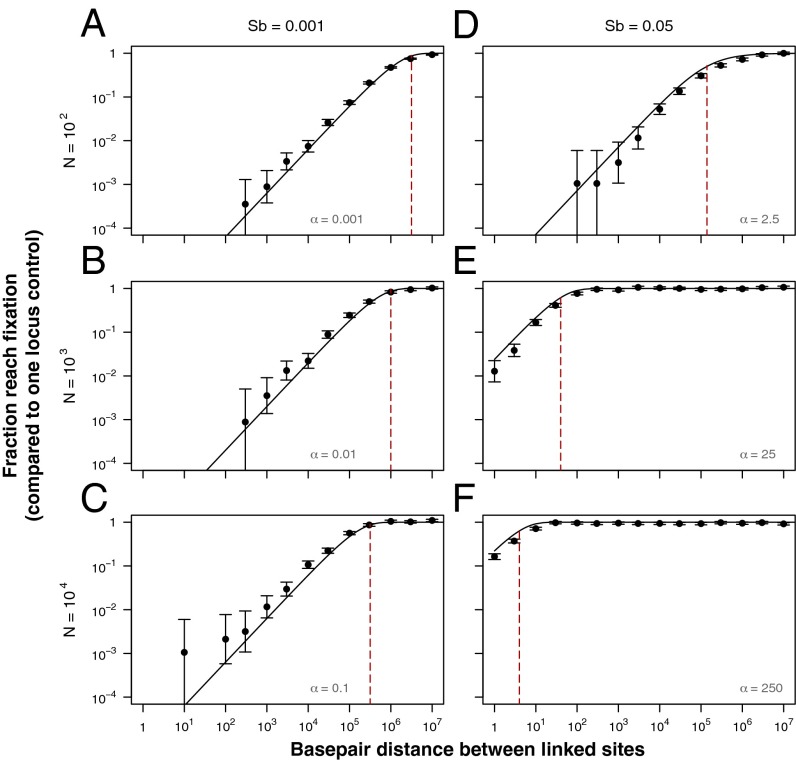
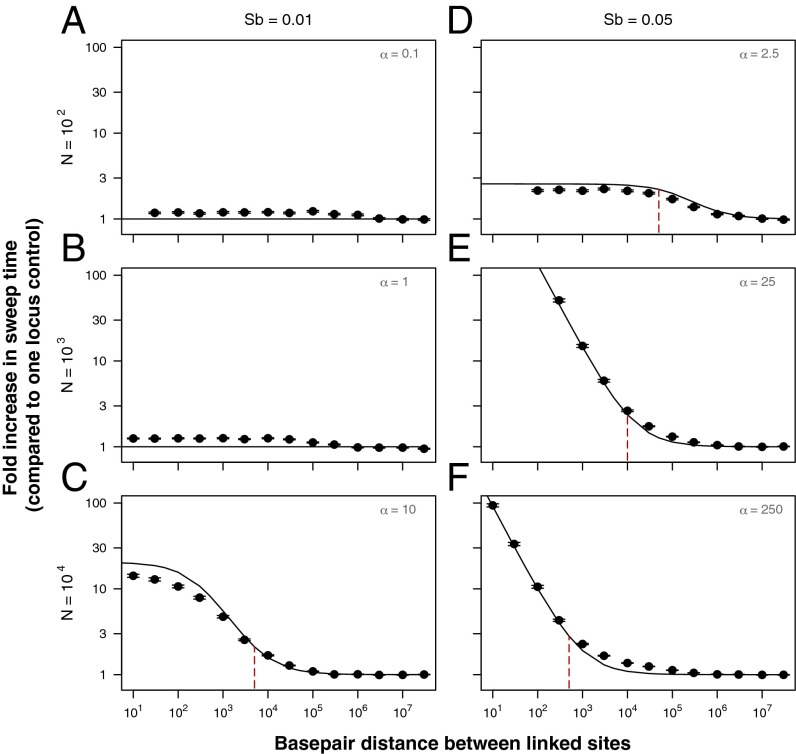
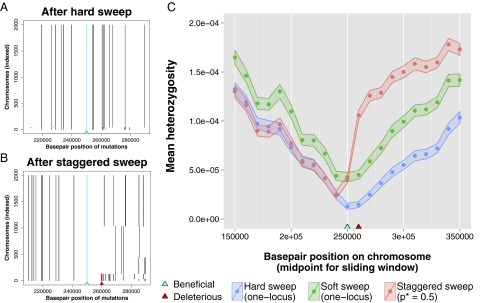
Recessive deleterious mutations are common, causing many genetic disorders in humans and producing inbreeding depression in the majority of sexually reproducing diploids. The abundance of recessive deleterious mutations in natural populations suggests they are likely to be present on a chromosome when a new adaptive mutation occurs, yet the dynamics of recessive deleterious hitchhikers and their impact on adaptation remains poorly understood. Here we model how a recessive deleterious mutation impacts the fate of a genetically linked dominant beneficial mutation. The frequency trajectory of the adaptive mutation in this case is dramatically altered and results in what we have termed a "staggered sweep." It is named for its three-phased trajectory: (i) Initially, the two linked mutations have a selective advantage while rare and will increase in frequency together, then (ii), at higher frequencies, the recessive hitchhiker is exposed to selection and can cause a balanced state via heterozygote advantage (the staggered phase), and (iii) finally, if recombination unlinks the two mutations, then the beneficial mutation can complete the sweep to fixation. Using both analytics and simulations, we show that strongly deleterious recessive mutations can substantially decrease the probability of fixation for nearby beneficial mutations, thus creating zones in the genome where adaptation is suppressed. These mutations can also significantly prolong the number of generations a beneficial mutation takes to sweep to fixation, and cause the genomic signature of selection to resemble that of soft or partial sweeps. We show that recessive deleterious variation could impact adaptation in humans and Drosophila.
Keywords: adaptation; hitchhiking; inbreeding depression; recessive; selective sweep.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Haldane JBS. A mathematical theory of natural and artificial selection, part V: Selection and mutation. Math Proc Cambridge Philos Soc. 1927;23(7):838−844.
-
- Charlesworth B. Adaptive evolution: The struggle for dominance. Curr Biol. 1998;8(14):R502–R504. - PubMed
-
- Fisher RA. The possible modification of the response of the wild type to recurrent mutations. Am Nat. 1928;62(1):115–116.
-
- Wright S. Physiological and evolutionary theories of dominance. Am Nat. 1934;68(714):24–53.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
