

DNA Sequence-Specific Binding of CENP-B Enhances the Fidelity of Human Centromere Function
- PMID: 25942623
- PMCID: PMC4421092
- DOI: 10.1016/j.devcel.2015.03.020
DNA Sequence-Specific Binding of CENP-B Enhances the Fidelity of Human Centromere Function
Abstract
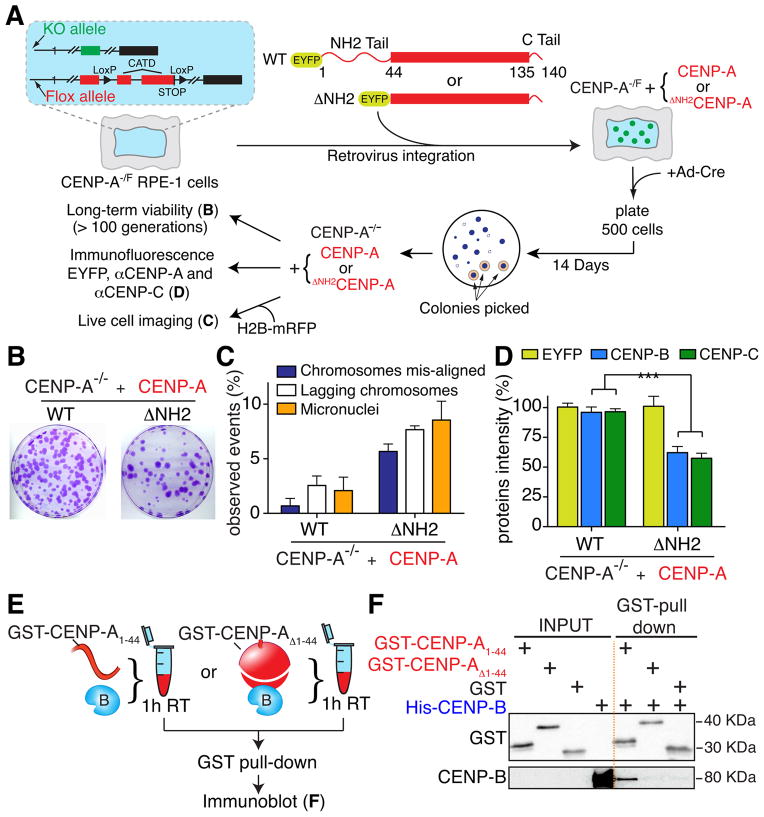
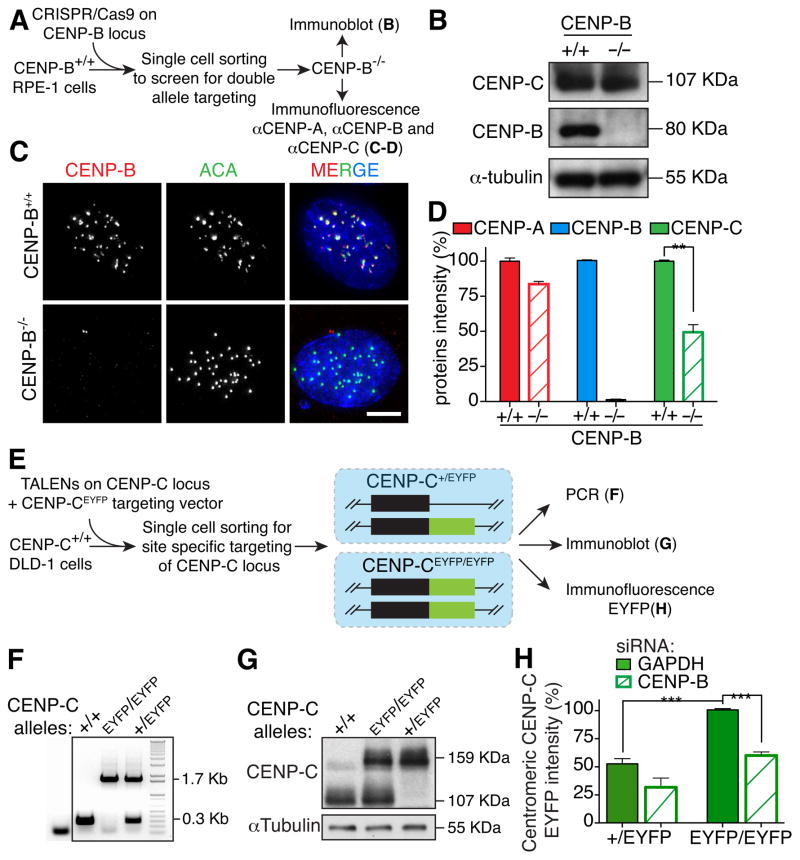
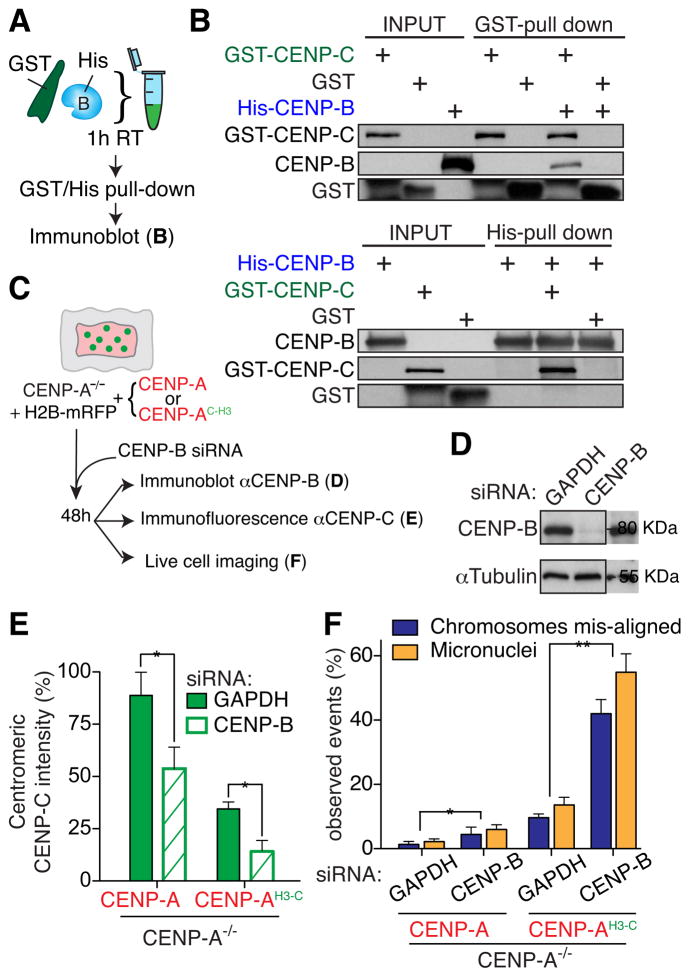
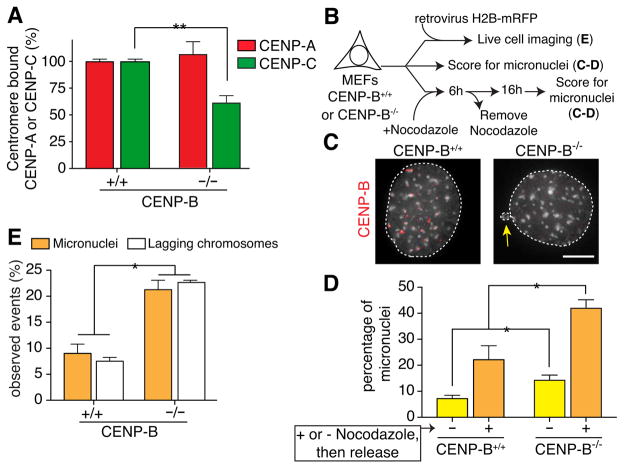
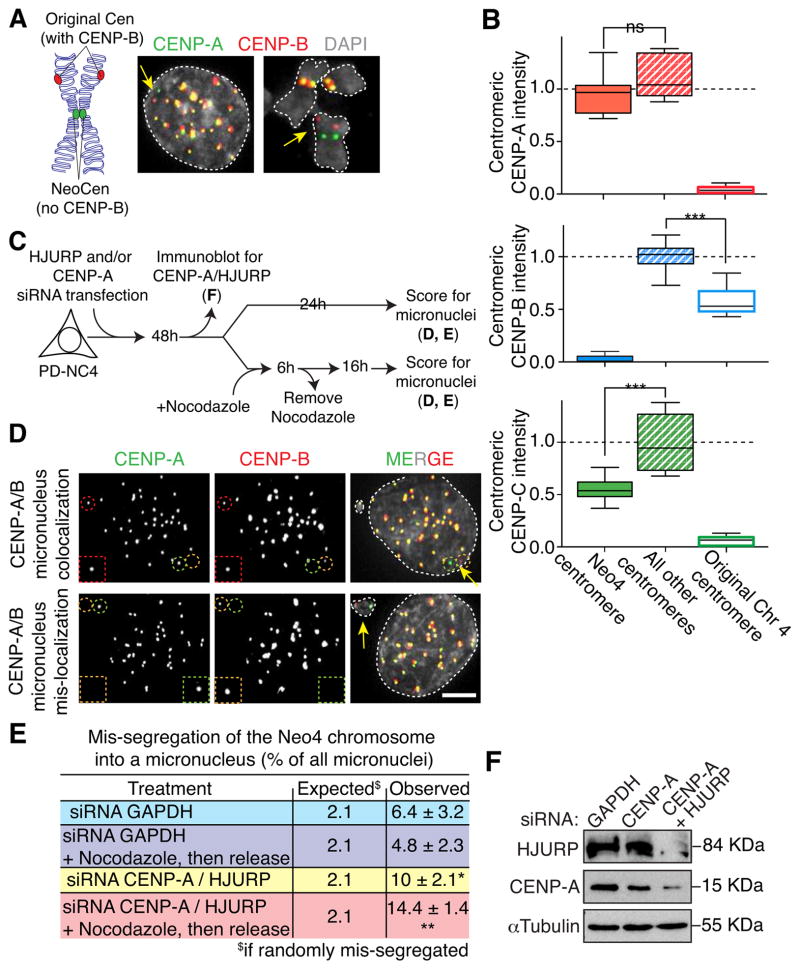
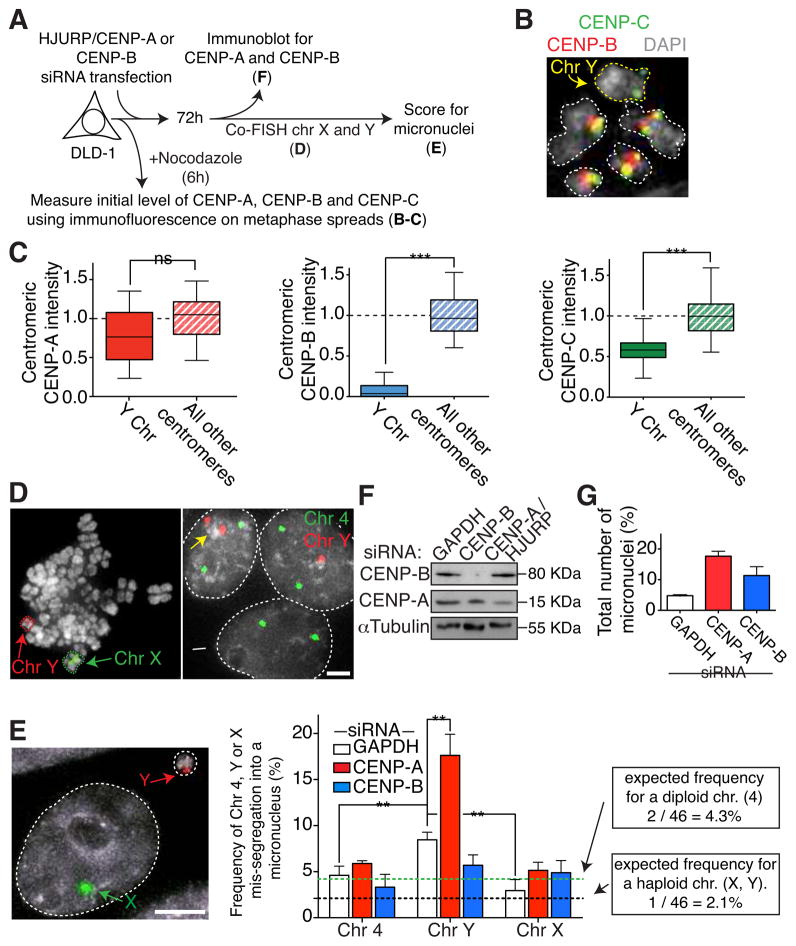
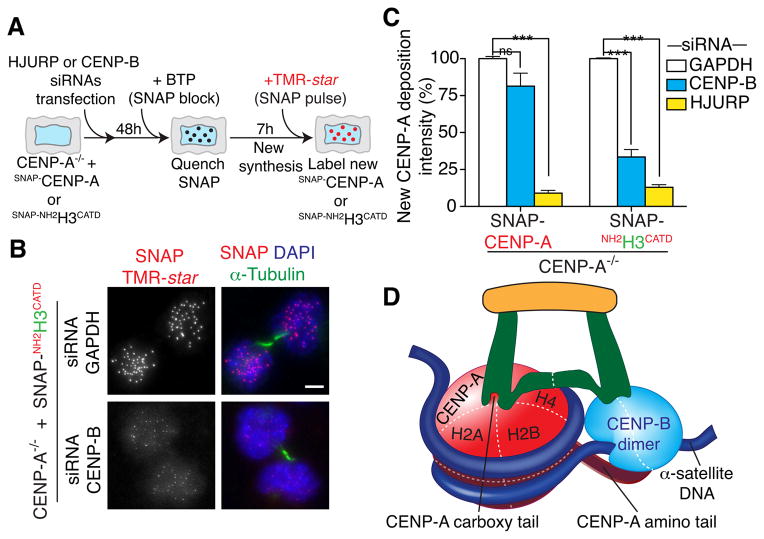
Human centromeres are specified by a stably inherited epigenetic mark that maintains centromere position and function through a two-step mechanism relying on self-templating centromeric chromatin assembled with the histone H3 variant CENP-A, followed by CENP-A-dependent nucleation of kinetochore assembly. Nevertheless, natural human centromeres are positioned within specific megabase chromosomal regions containing α-satellite DNA repeats, which contain binding sites for the DNA sequence-specific binding protein CENP-B. We now demonstrate that CENP-B directly binds both CENP-A's amino-terminal tail and CENP-C, a key nucleator of kinetochore assembly. DNA sequence-dependent binding of CENP-B within α-satellite repeats is required to stabilize optimal centromeric levels of CENP-C. Chromosomes bearing centromeres without bound CENP-B, including the human Y chromosome, are shown to mis-segregate in cells at rates several-fold higher than chromosomes with CENP-B-containing centromeres. These data demonstrate a DNA sequence-specific enhancement by CENP-B of the fidelity of epigenetically defined human centromere function.
Copyright © 2015 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
