

Prdx4 is a compartment-specific H2O2 sensor that regulates neurogenesis by controlling surface expression of GDE2
- PMID: 25943695
- PMCID: PMC4432624
- DOI: 10.1038/ncomms8006
Prdx4 is a compartment-specific H2O2 sensor that regulates neurogenesis by controlling surface expression of GDE2
Abstract
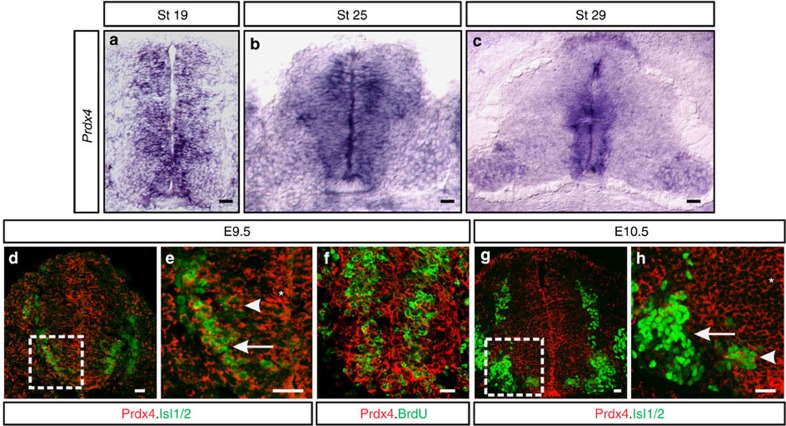
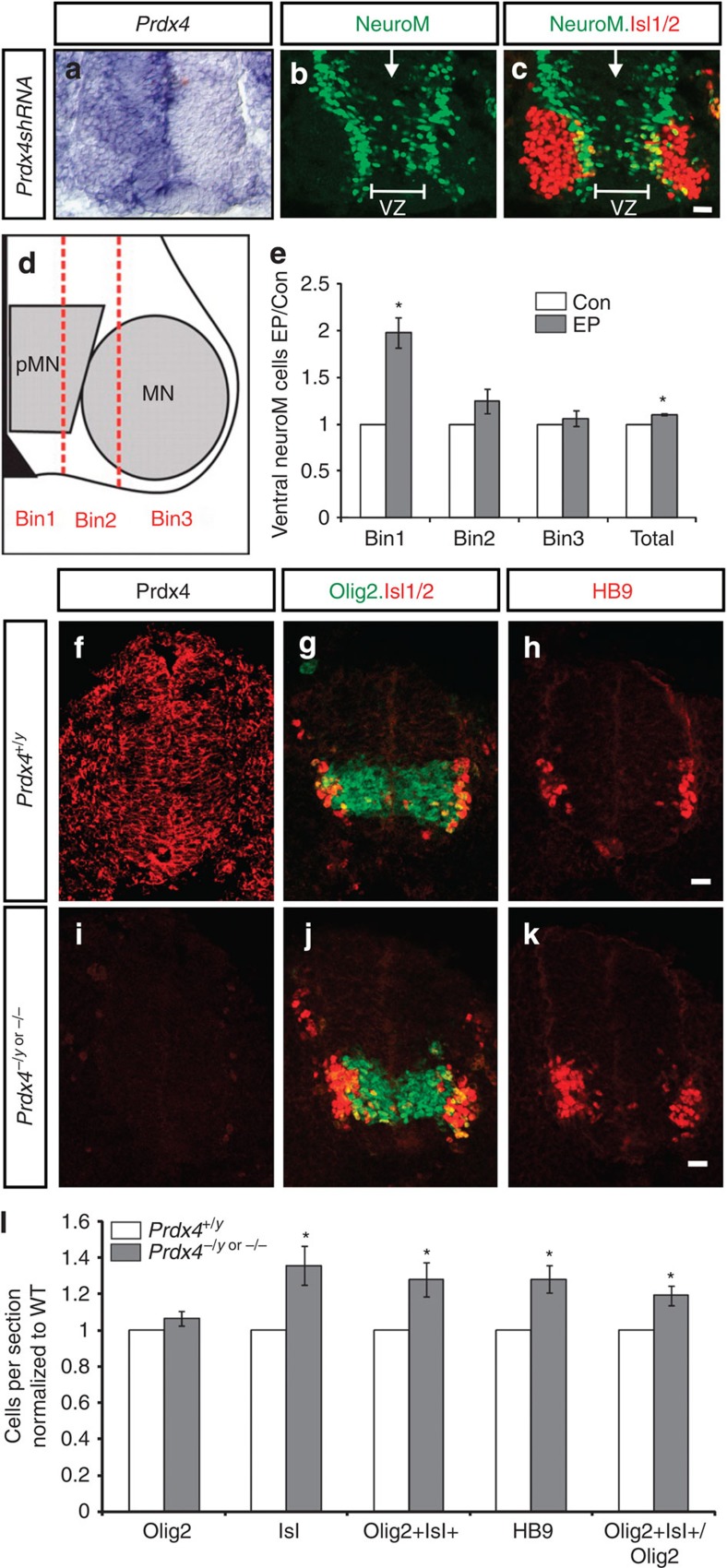
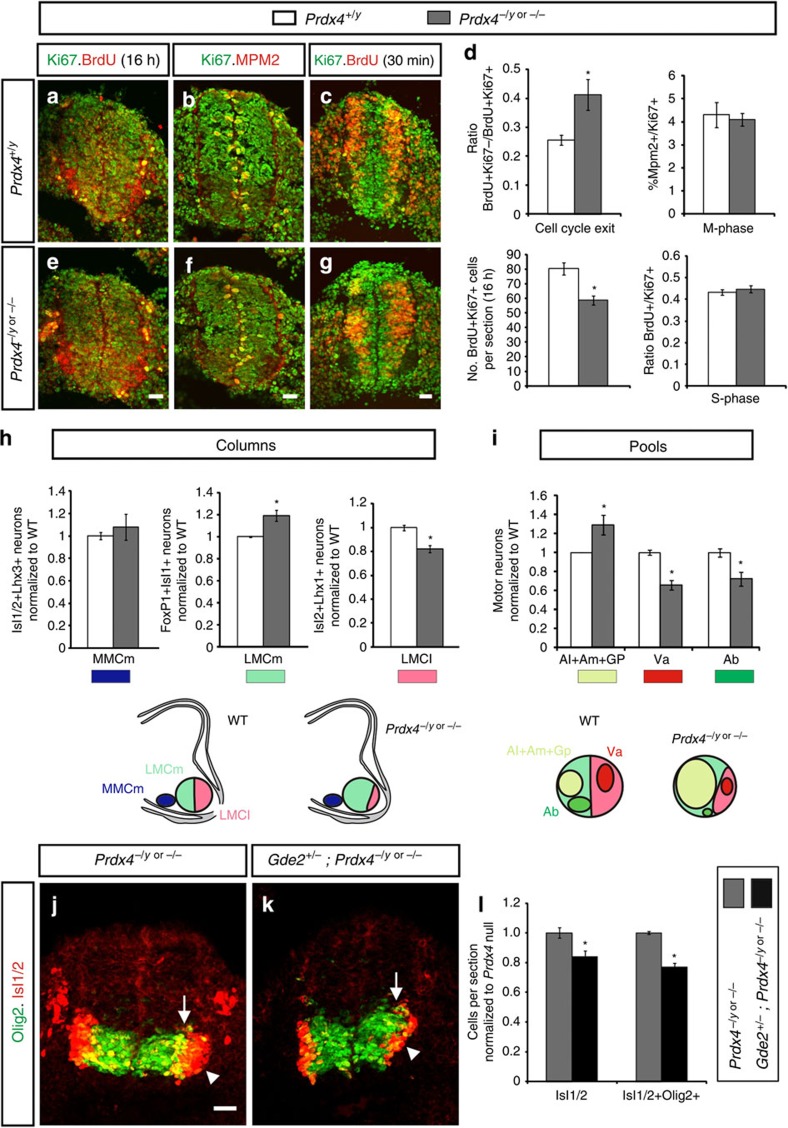
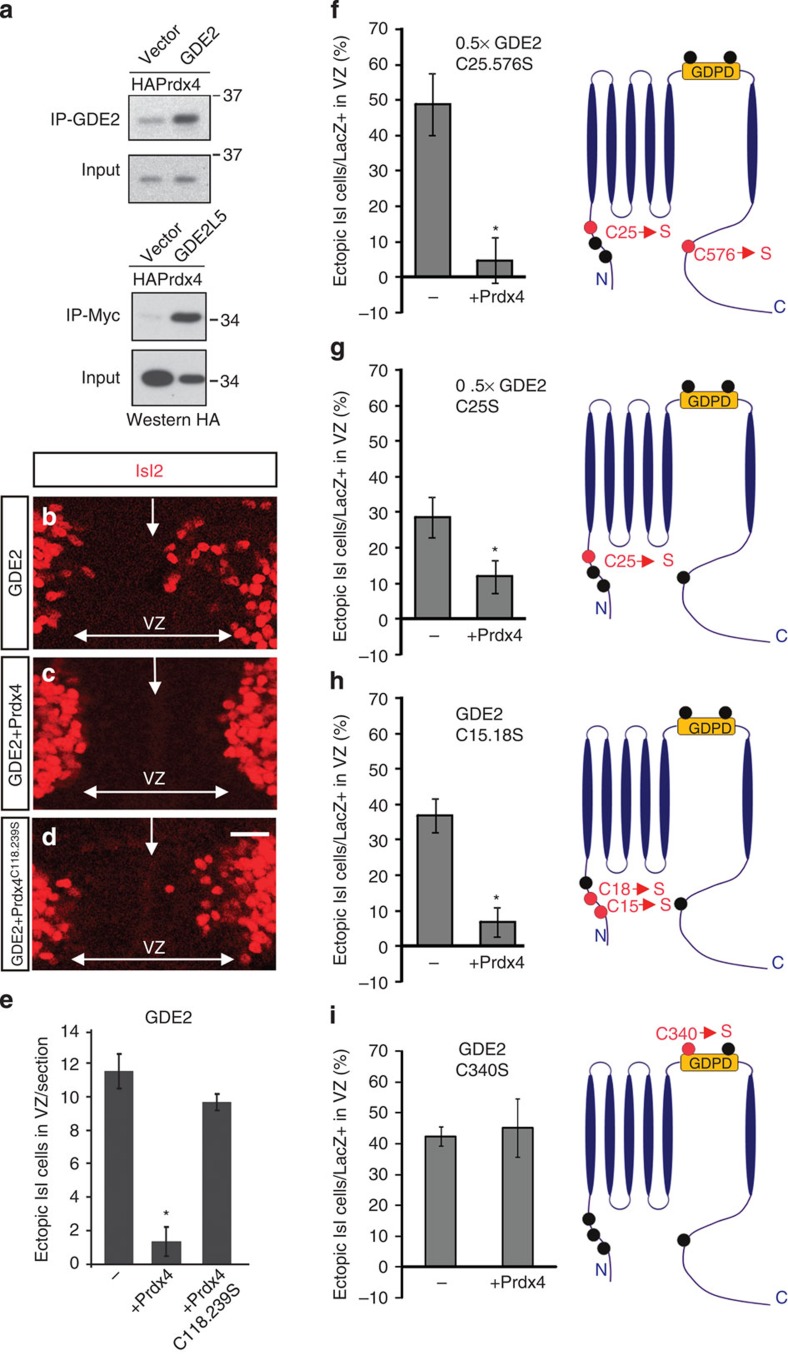
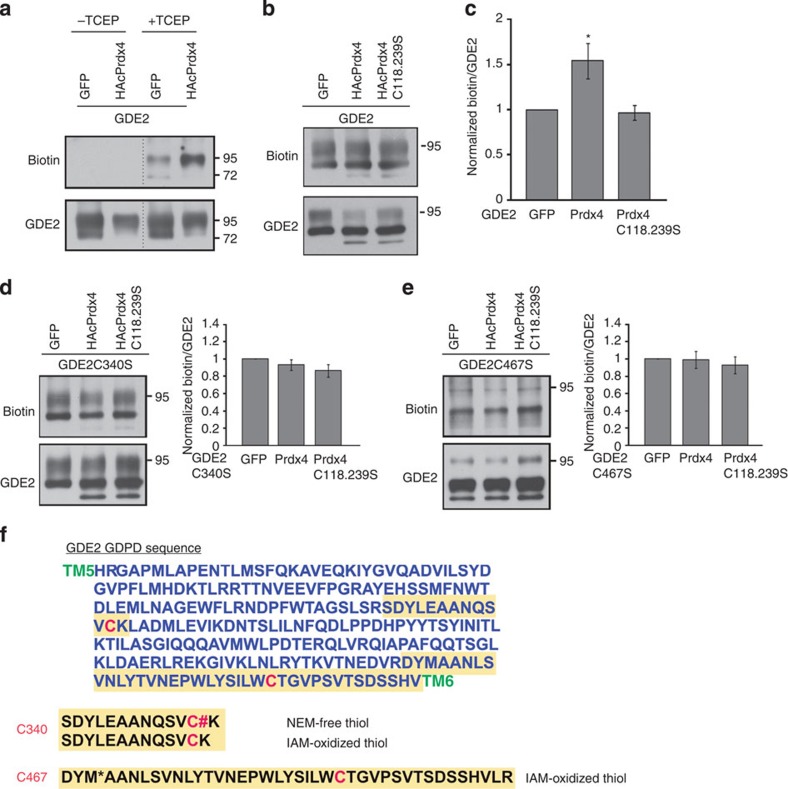
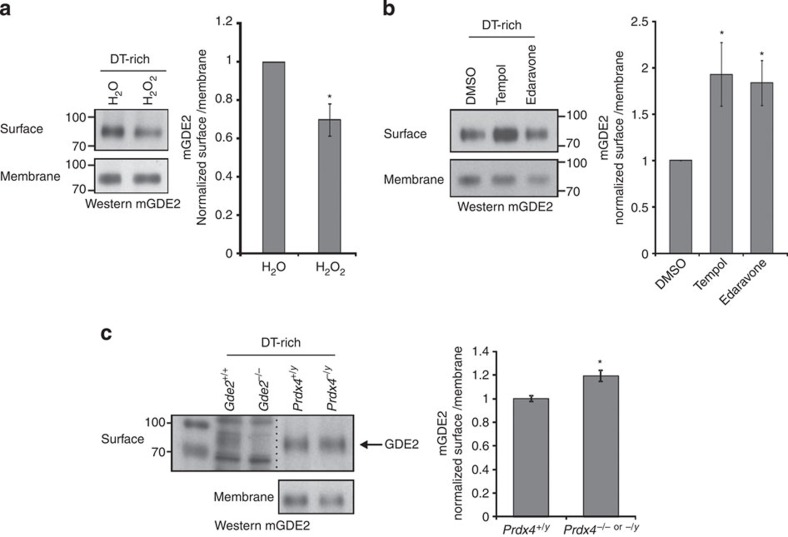
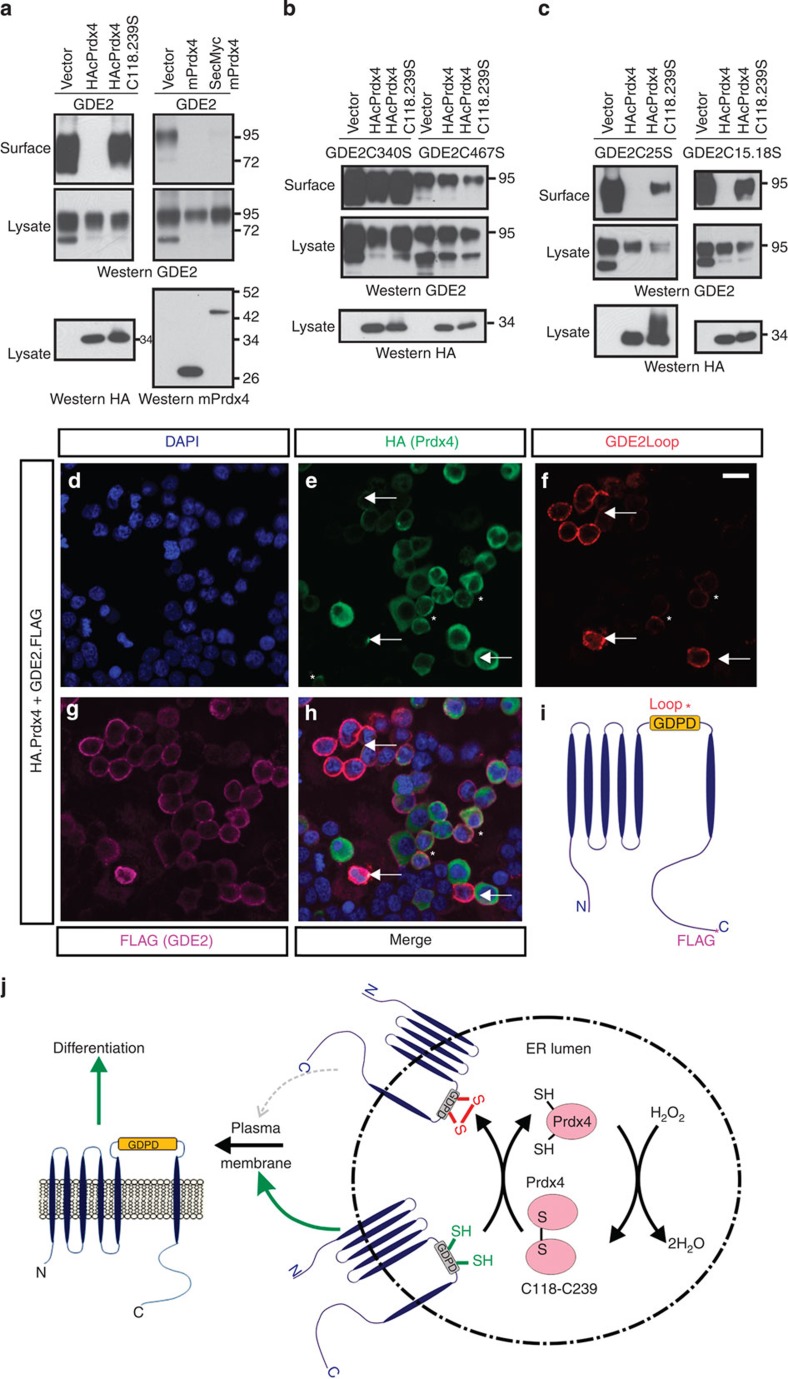
Neural progenitors and terminally differentiated neurons show distinct redox profiles, suggesting that coupled-redox cascades regulate the initiation and progression of neuronal differentiation. Discrete cellular compartments have different redox environments and how they contribute to differentiation is unclear. Here we show that Prdx4, an endoplasmic reticulum (ER) enzyme that metabolizes H2O2, acts as a tunable regulator of neurogenesis via its compartmentalized thiol-oxidative function. Prdx4 ablation causes premature motor neuron differentiation and progenitor depletion, leading to imbalances in subtype-specific motor neurons. GDE2, a six-transmembrane protein that induces differentiation by downregulating Notch signalling through surface cleavage of GPI-anchored proteins, is targeted by Prdx4 oxidative activity. Prdx4 dimers generated by H2O2 metabolism oxidize two cysteine residues within the GDE2 enzymatic domain, which blocks GDE2 trafficking to the plasma membrane and prevents GDE2 neurogeneic function. Thus, Prdx4 oxidative activity acts as a sensor to directly couple neuronal differentiation with redox environments in the ER.
Figures
Similar articles
-
The antioxidant enzyme Prdx1 controls neuronal differentiation by thiol-redox-dependent activation of GDE2.Cell. 2009 Sep 18;138(6):1209-21. doi: 10.1016/j.cell.2009.06.042. Cell. 2009. PMID: 19766572 Free PMC article.
-
GDE2 promotes neurogenesis by glycosylphosphatidylinositol-anchor cleavage of RECK.Science. 2013 Jan 18;339(6117):324-8. doi: 10.1126/science.1231921. Science. 2013. PMID: 23329048 Free PMC article.
-
GDE2 is essential for neuronal survival in the postnatal mammalian spinal cord.Mol Neurodegener. 2017 Jan 19;12(1):8. doi: 10.1186/s13024-017-0148-1. Mol Neurodegener. 2017. PMID: 28103900 Free PMC article.
-
PRDX4, an endoplasmic reticulum-localized peroxiredoxin at the crossroads between enzymatic oxidative protein folding and nonenzymatic protein oxidation.Antioxid Redox Signal. 2013 May 1;18(13):1666-74. doi: 10.1089/ars.2012.4966. Epub 2012 Nov 6. Antioxid Redox Signal. 2013. PMID: 23025503 Review.
-
The Conundrum of Hydrogen Peroxide Signaling and the Emerging Role of Peroxiredoxins as Redox Relay Hubs.Antioxid Redox Signal. 2018 Mar 1;28(7):558-573. doi: 10.1089/ars.2017.7162. Epub 2017 Jul 17. Antioxid Redox Signal. 2018. PMID: 28587525 Review.
Cited by
-
Hydrogen Peroxide and Redox Regulation of Developments.Antioxidants (Basel). 2018 Nov 6;7(11):159. doi: 10.3390/antiox7110159. Antioxidants (Basel). 2018. PMID: 30404180 Free PMC article. Review.
-
Expression and prognostic roles of PRDXs gene family in hepatocellular carcinoma.J Transl Med. 2021 Mar 26;19(1):126. doi: 10.1186/s12967-021-02792-8. J Transl Med. 2021. PMID: 33771165 Free PMC article.
-
Genome-wide association analysis of egg production performance in chickens across the whole laying period.BMC Genet. 2019 Aug 14;20(1):67. doi: 10.1186/s12863-019-0771-7. BMC Genet. 2019. PMID: 31412760 Free PMC article.
-
TiO₂-Based Nanoheterostructures for Promoting Gas Sensitivity Performance: Designs, Developments, and Prospects.Sensors (Basel). 2017 Aug 27;17(9):1971. doi: 10.3390/s17091971. Sensors (Basel). 2017. PMID: 28846621 Free PMC article. Review.
-
Microglia antioxidant systems and redox signalling.Br J Pharmacol. 2017 Jun;174(12):1719-1732. doi: 10.1111/bph.13426. Epub 2016 Mar 3. Br J Pharmacol. 2017. PMID: 26754582 Free PMC article. Review.
References
-
- Schafer F. Q. & Buettner G. R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 30, 1191–1212 (2001). - PubMed
-
- Putker M., Vos H. R. & Dansen T. B. Intermolecular disulfide-dependent redox signalling. Biochem. Soc. Trans. 42, 971–978 (2014). - PubMed
-
- Holmström K. M. & Finkel T. Cellular mechanisms and physiological consequences of redox-dependent signalling. Nat. Rev. Mol. Cell. Biol. 15, 411–421 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
