

Activity-dependent, homeostatic regulation of neurotransmitter release from auditory nerve fibers
- PMID: 25944933
- PMCID: PMC4443347
- DOI: 10.1073/pnas.1420885112
Activity-dependent, homeostatic regulation of neurotransmitter release from auditory nerve fibers
Abstract
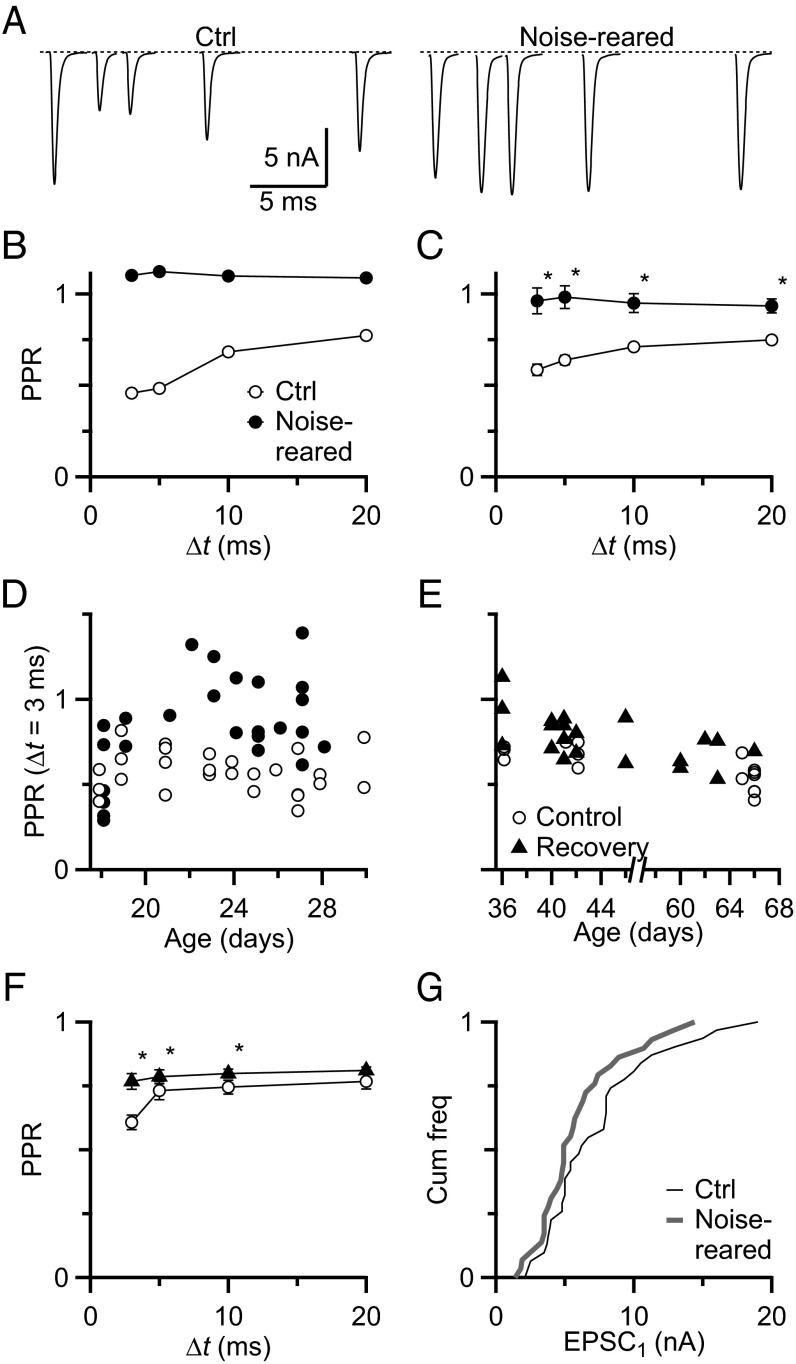
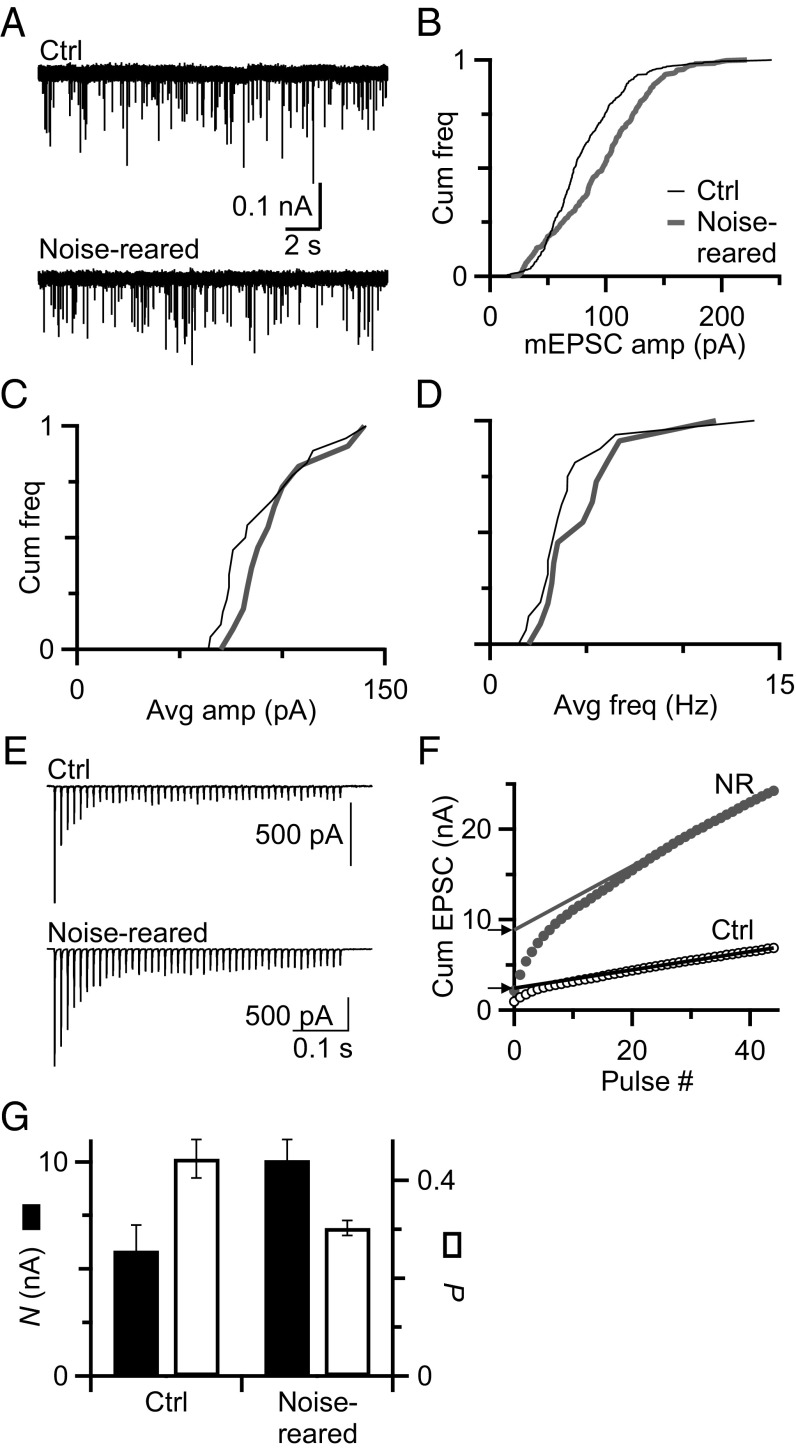
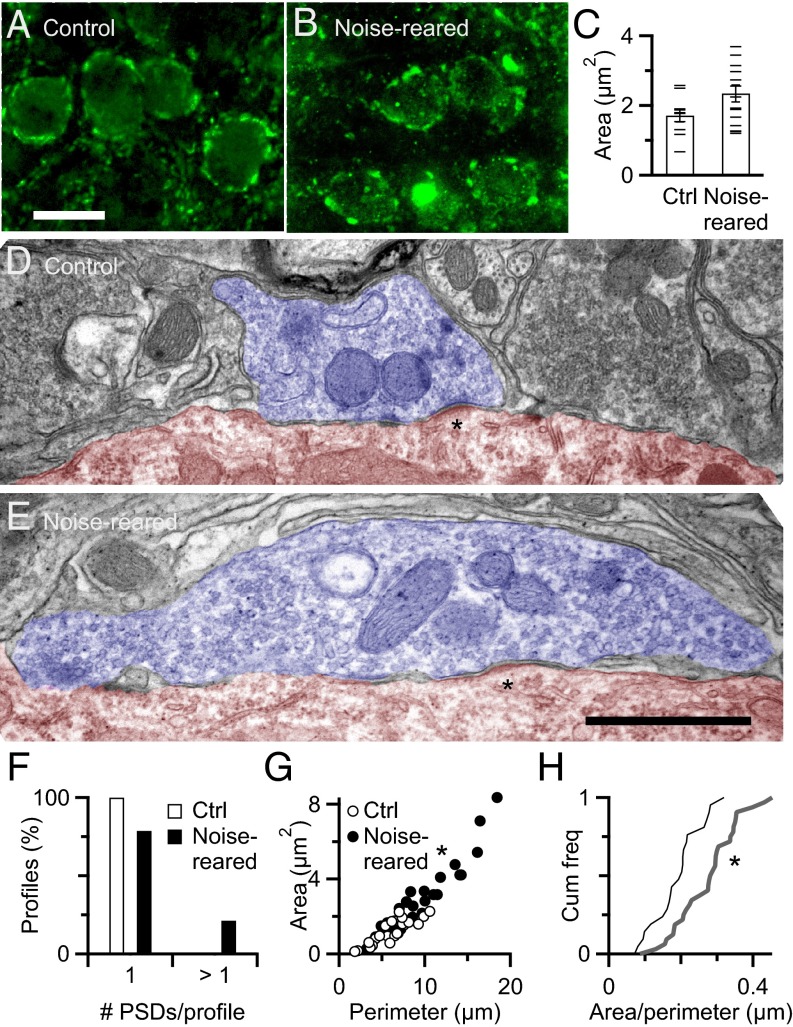
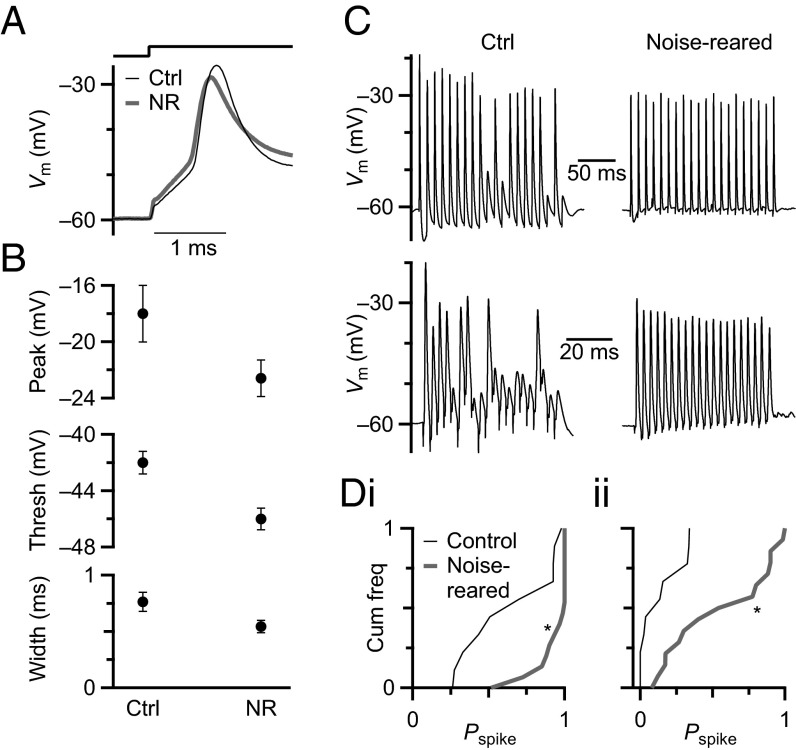
Information processing in the brain requires reliable synaptic transmission. High reliability at specialized auditory nerve synapses in the cochlear nucleus results from many release sites (N), high probability of neurotransmitter release (Pr), and large quantal size (Q). However, high Pr also causes auditory nerve synapses to depress strongly when activated at normal rates for a prolonged period, which reduces fidelity. We studied how synapses are influenced by prolonged activity by exposing mice to constant, nondamaging noise and found that auditory nerve synapses changed to facilitating, reflecting low Pr. For mice returned to quiet, synapses recovered to normal depression, suggesting that these changes are a homeostatic response to activity. Two additional properties, Q and average excitatory postsynaptic current (EPSC) amplitude, were unaffected by noise rearing, suggesting that the number of release sites (N) must increase to compensate for decreased Pr. These changes in N and Pr were confirmed physiologically using the integration method. Furthermore, consistent with increased N, endbulbs in noise-reared animals had larger VGlut1-positive puncta, larger profiles in electron micrographs, and more release sites per profile. In current-clamp recordings, noise-reared BCs had greater spike fidelity even during high rates of synaptic activity. Thus, auditory nerve synapses regulate excitability through an activity-dependent, homeostatic mechanism, which could have major effects on all downstream processing. Our results also suggest that noise-exposed bushy cells would remain hyperexcitable for a period after returning to normal quiet conditions, which could have perceptual consequences.
Keywords: cochlear nucleus; homeostasis; release probability.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Turrigiano GG, Leslie KR, Desai NS, Rutherford LC, Nelson SB. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature. 1998;391(6670):892–896. - PubMed
-
- Turrigiano GG, Nelson SB. Homeostatic plasticity in the developing nervous system. Nat Rev Neurosci. 2004;5(2):97–107. - PubMed
-
- O’Brien RJ, et al. Activity-dependent modulation of synaptic AMPA receptor accumulation. Neuron. 1998;21(5):1067–1078. - PubMed
-
- Keck T, et al. Synaptic scaling and homeostatic plasticity in the mouse visual cortex in vivo. Neuron. 2013;80(2):327–334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
