

Assembly of a comprehensive regulatory network for the mammalian circadian clock: a bioinformatics approach
- PMID: 25945798
- PMCID: PMC4422523
- DOI: 10.1371/journal.pone.0126283
Assembly of a comprehensive regulatory network for the mammalian circadian clock: a bioinformatics approach
Abstract
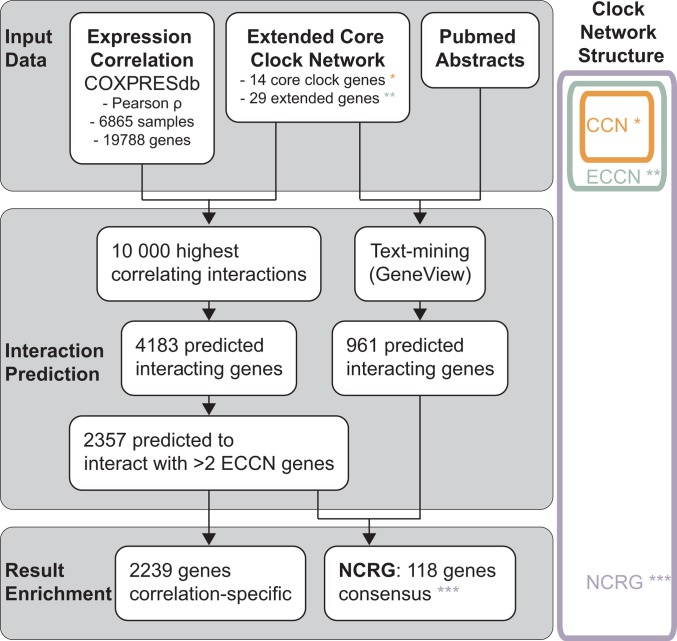
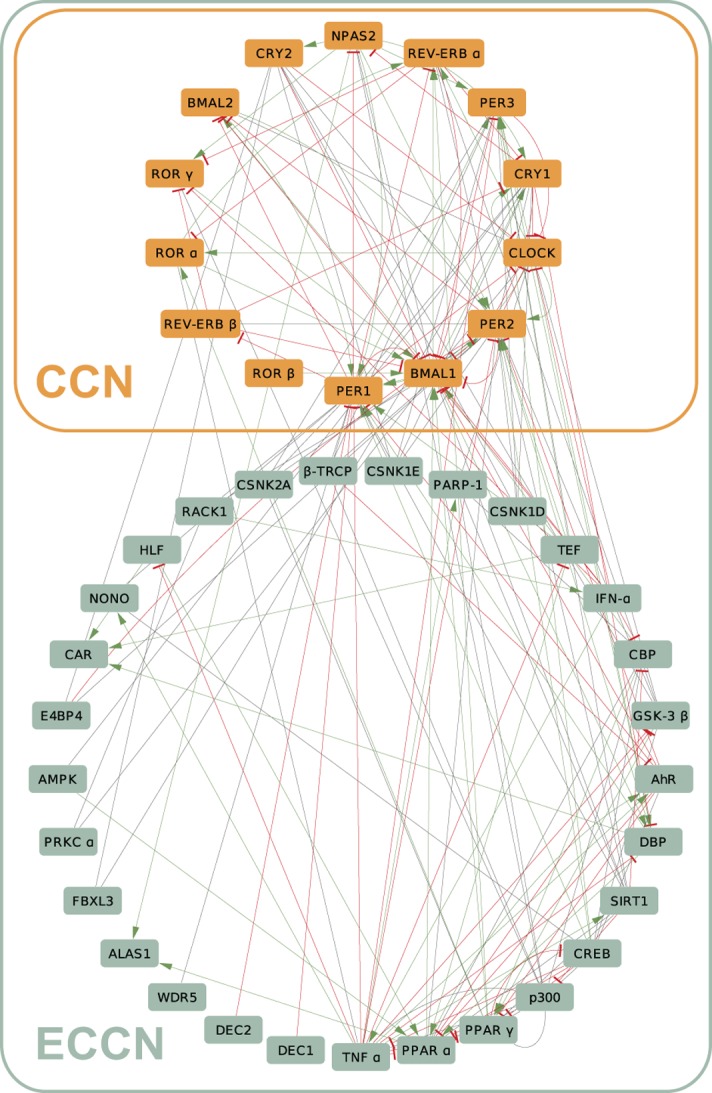
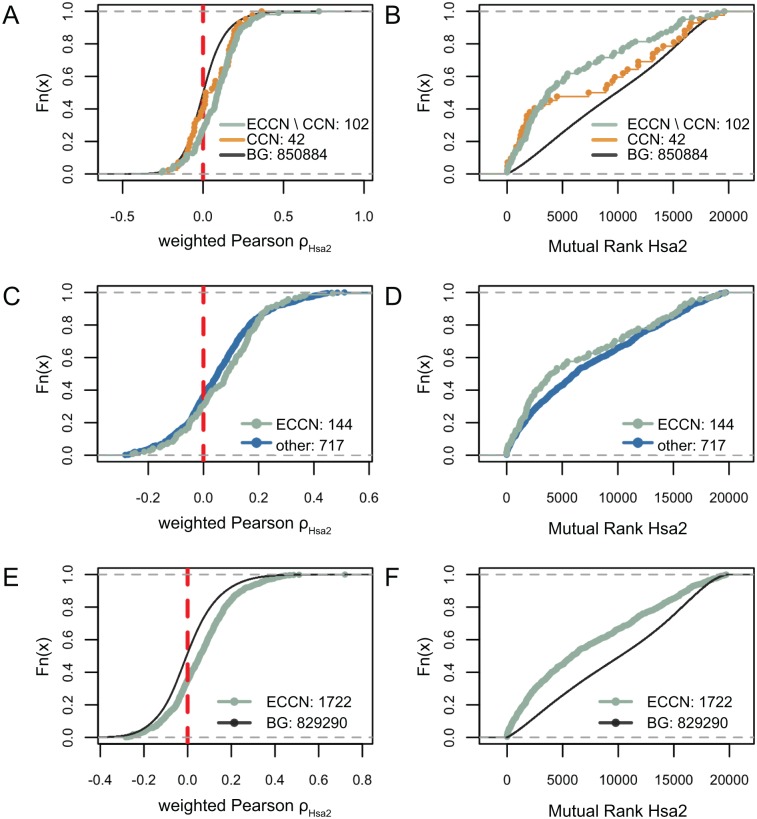
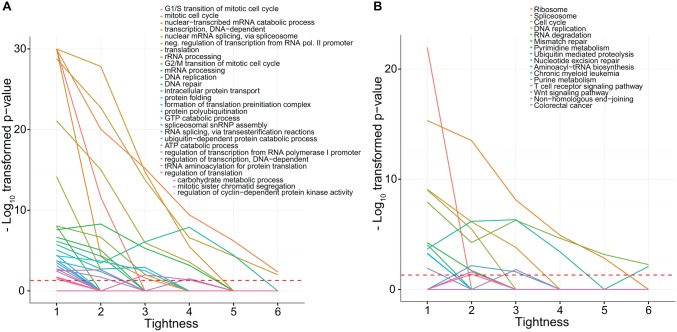
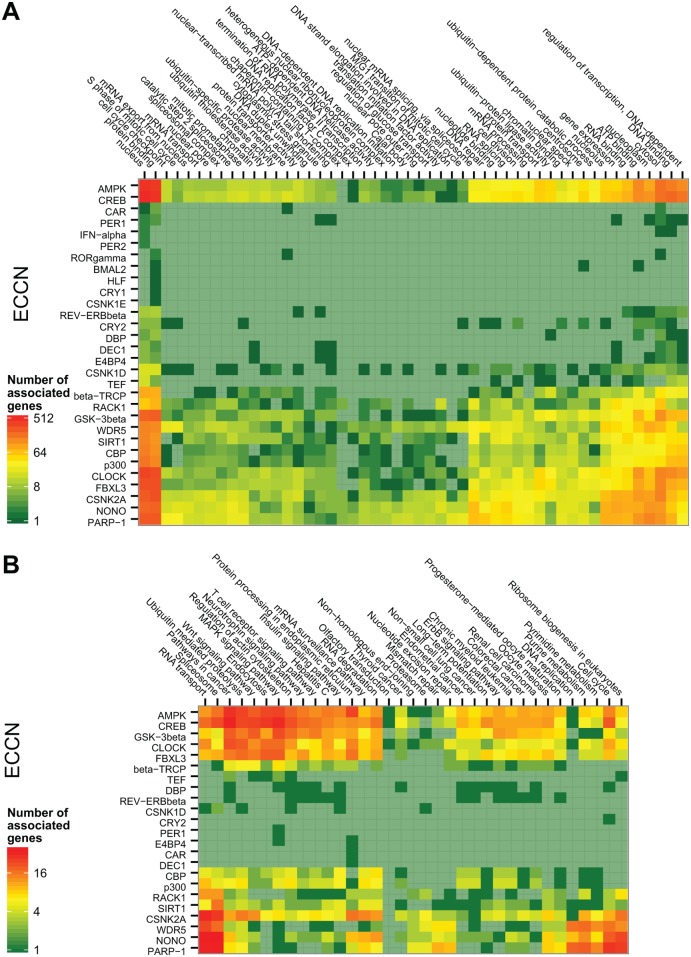
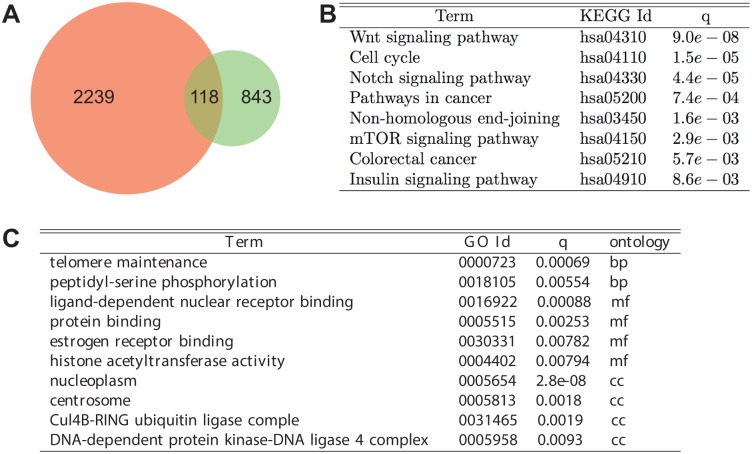
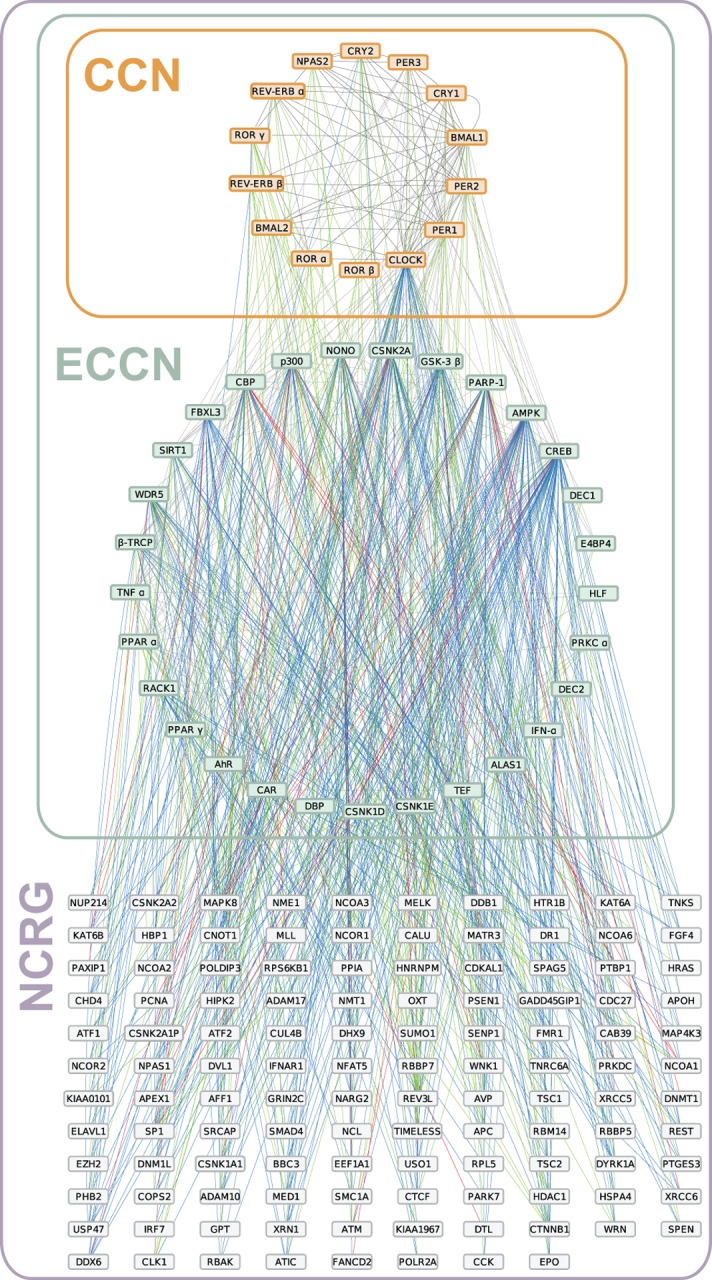
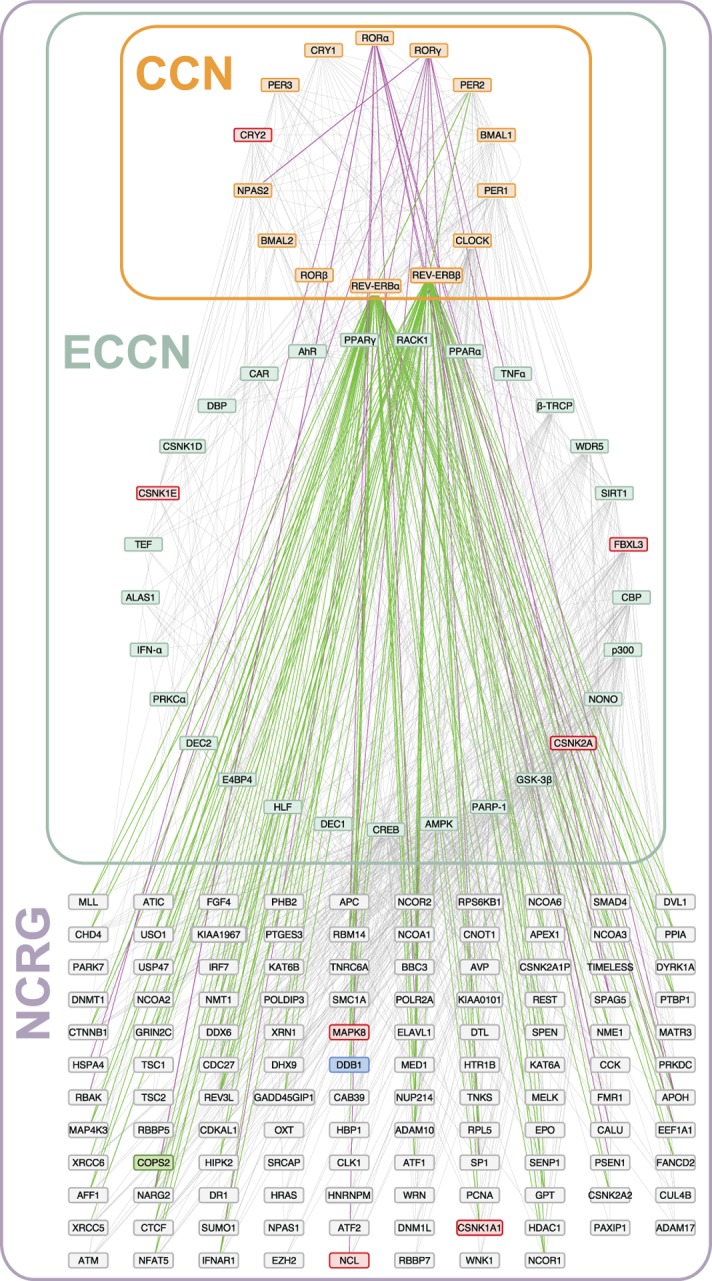
By regulating the timing of cellular processes, the circadian clock provides a way to adapt physiology and behaviour to the geophysical time. In mammals, a light-entrainable master clock located in the suprachiasmatic nucleus (SCN) controls peripheral clocks that are present in virtually every body cell. Defective circadian timing is associated with several pathologies such as cancer and metabolic and sleep disorders. To better understand the circadian regulation of cellular processes, we developed a bioinformatics pipeline encompassing the analysis of high-throughput data sets and the exploitation of published knowledge by text-mining. We identified 118 novel potential clock-regulated genes and integrated them into an existing high-quality circadian network, generating the to-date most comprehensive network of circadian regulated genes (NCRG). To validate particular elements in our network, we assessed publicly available ChIP-seq data for BMAL1, REV-ERBα/β and RORα/γ proteins and found strong evidence for circadian regulation of Elavl1, Nme1, Dhx6, Med1 and Rbbp7 all of which are involved in the regulation of tumourigenesis. Furthermore, we identified Ncl and Ddx6, as targets of RORγ and REV-ERBα, β, respectively. Most interestingly, these genes were also reported to be involved in miRNA regulation; in particular, NCL regulates several miRNAs, all involved in cancer aggressiveness. Thus, NCL represents a novel potential link via which the circadian clock, and specifically RORγ, regulates the expression of miRNAs, with particular consequences in breast cancer progression. Our findings bring us one step forward towards a mechanistic understanding of mammalian circadian regulation, and provide further evidence of the influence of circadian deregulation in cancer.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
