

Sequential conformational changes in the morbillivirus attachment protein initiate the membrane fusion process
- PMID: 25946112
- PMCID: PMC4422687
- DOI: 10.1371/journal.ppat.1004880
Sequential conformational changes in the morbillivirus attachment protein initiate the membrane fusion process
Abstract
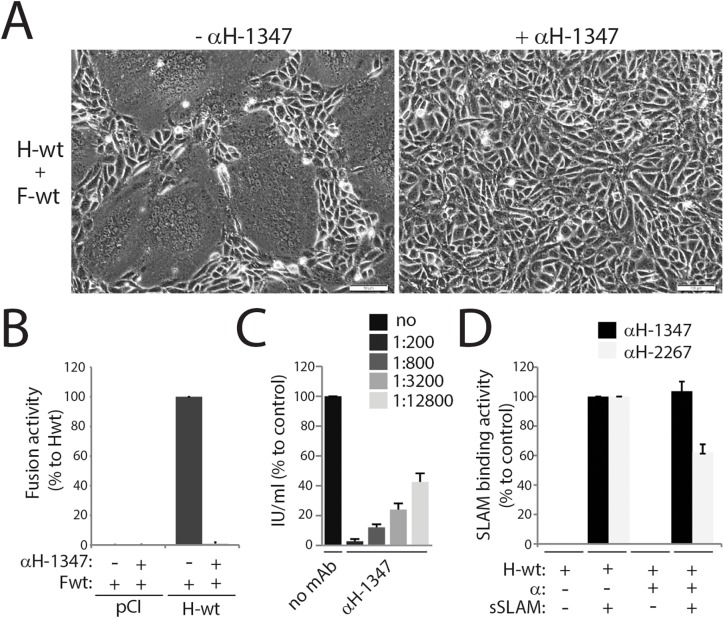
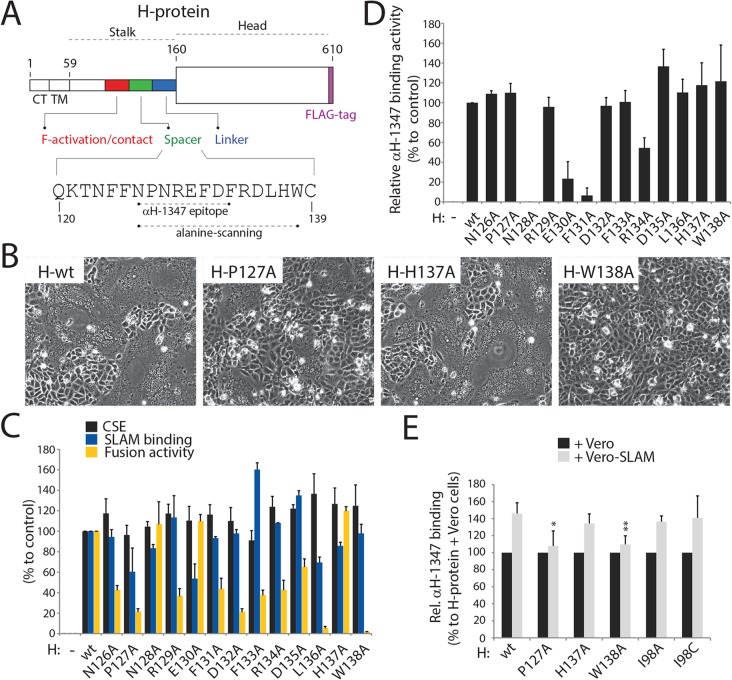
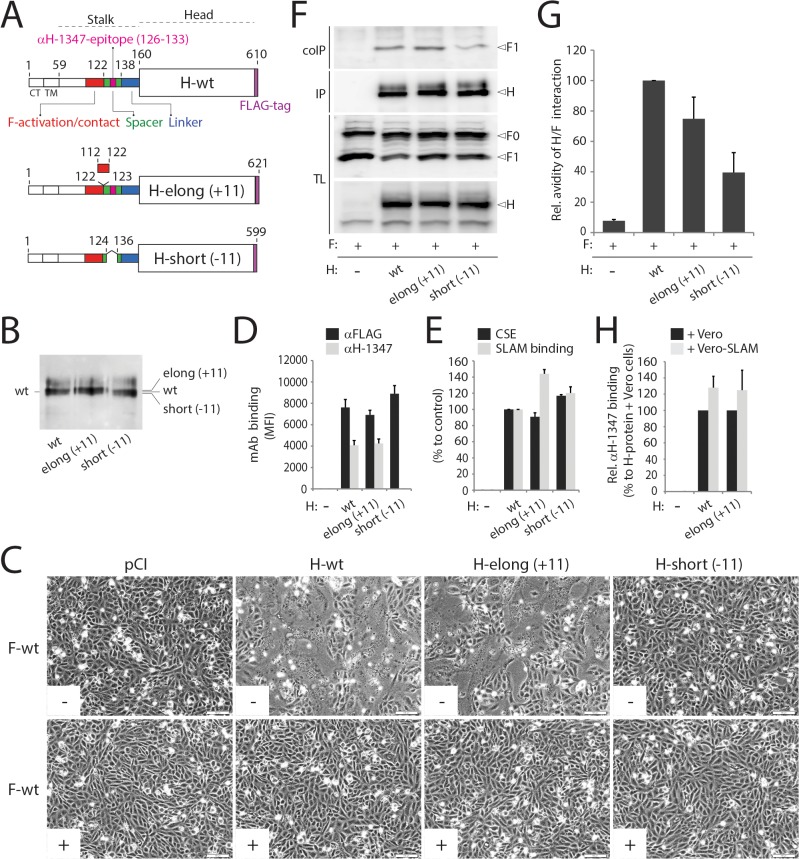
Despite large vaccination campaigns, measles virus (MeV) and canine distemper virus (CDV) cause major morbidity and mortality in humans and animals, respectively. The MeV and CDV cell entry system relies on two interacting envelope glycoproteins: the attachment protein (H), consisting of stalk and head domains, co-operates with the fusion protein (F) to mediate membrane fusion. However, how receptor-binding by the H-protein leads to F-triggering is not fully understood. Here, we report that an anti-CDV-H monoclonal antibody (mAb-1347), which targets the linear H-stalk segment 126-133, potently inhibits membrane fusion without interfering with H receptor-binding or F-interaction. Rather, mAb-1347 blocked the F-triggering function of H-proteins regardless of the presence or absence of the head domains. Remarkably, mAb-1347 binding to headless CDV H, as well as standard and engineered bioactive stalk-elongated CDV H-constructs treated with cells expressing the SLAM receptor, was enhanced. Despite proper cell surface expression, fusion promotion by most H-stalk mutants harboring alanine substitutions in the 126-138 "spacer" section was substantially impaired, consistent with deficient receptor-induced mAb-1347 binding enhancement. However, a previously reported F-triggering defective H-I98A variant still exhibited the receptor-induced "head-stalk" rearrangement. Collectively, our data spotlight a distinct mechanism for morbillivirus membrane fusion activation: prior to receptor contact, at least one of the morbillivirus H-head domains interacts with the membrane-distal "spacer" domain in the H-stalk, leaving the F-binding site located further membrane-proximal in the stalk fully accessible. This "head-to-spacer" interaction conformationally stabilizes H in an auto-repressed state, which enables intracellular H-stalk/F engagement while preventing the inherent H-stalk's bioactivity that may prematurely activate F. Receptor-contact disrupts the "head-to-spacer" interaction, which subsequently "unlocks" the stalk, allowing it to rearrange and trigger F. Overall, our study reveals essential mechanistic requirements governing the activation of the morbillivirus membrane fusion cascade and spotlights the H-stalk "spacer" microdomain as a possible drug target for antiviral therapy.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
