

The Tyrosine Kinase c-Src Specifically Binds to the Active Integrin αIIbβ3 to Initiate Outside-in Signaling in Platelets
- PMID: 25947380
- PMCID: PMC4505490
- DOI: 10.1074/jbc.M115.648428
The Tyrosine Kinase c-Src Specifically Binds to the Active Integrin αIIbβ3 to Initiate Outside-in Signaling in Platelets
Abstract
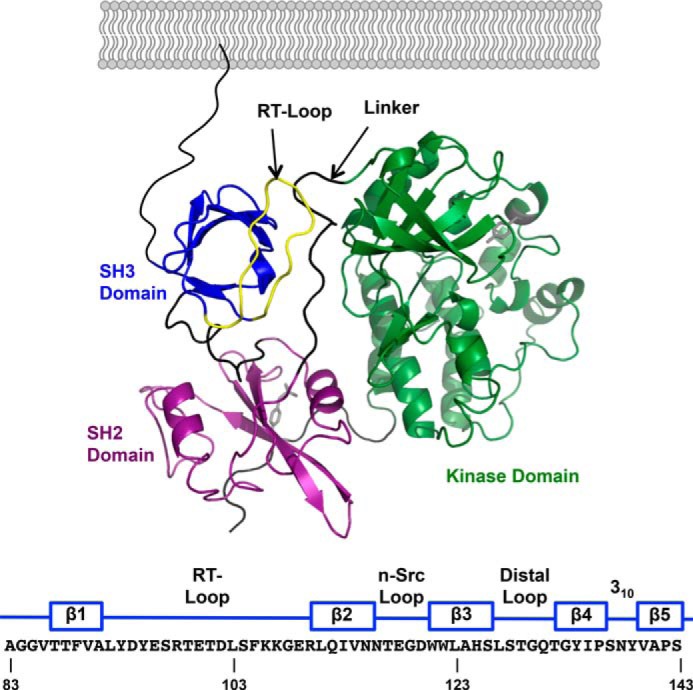
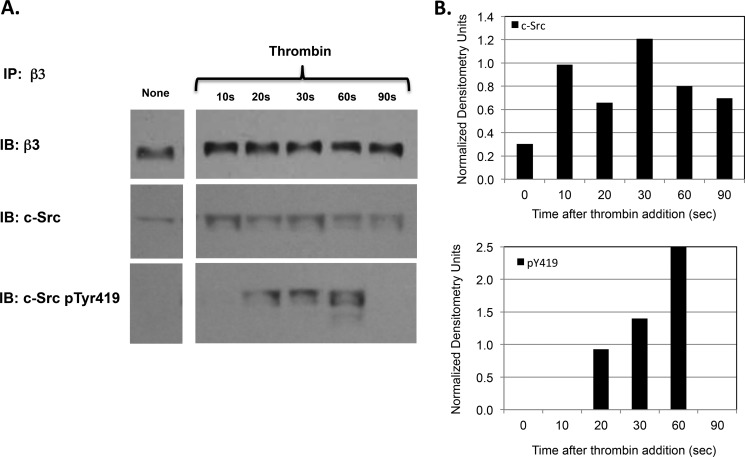
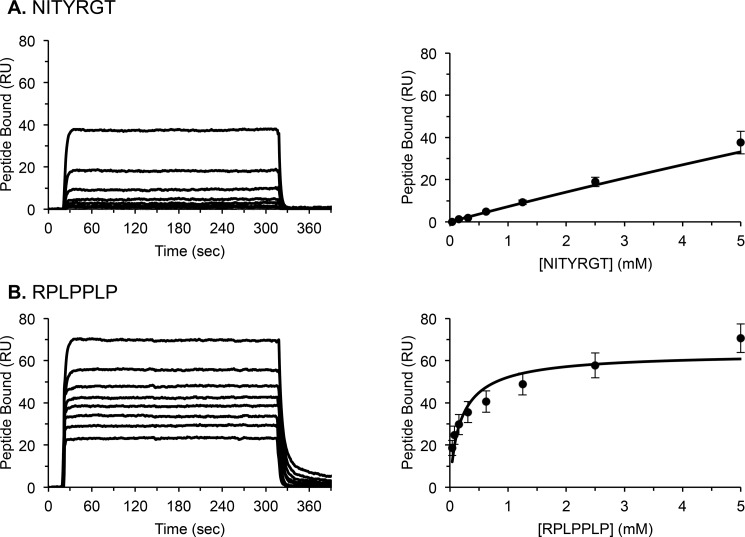
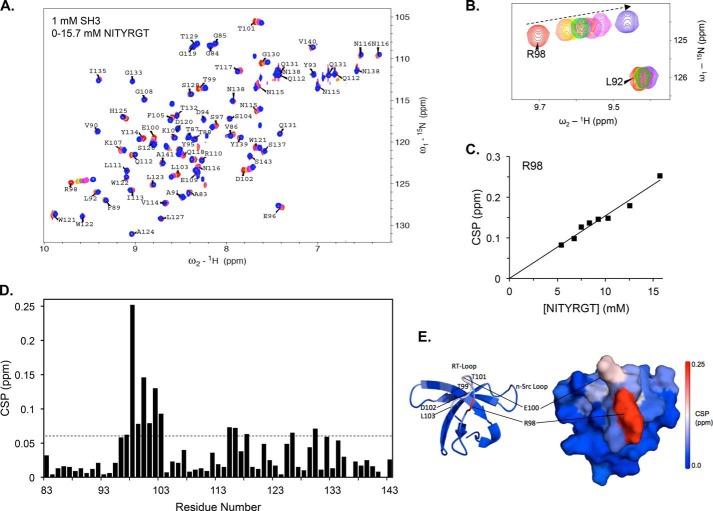
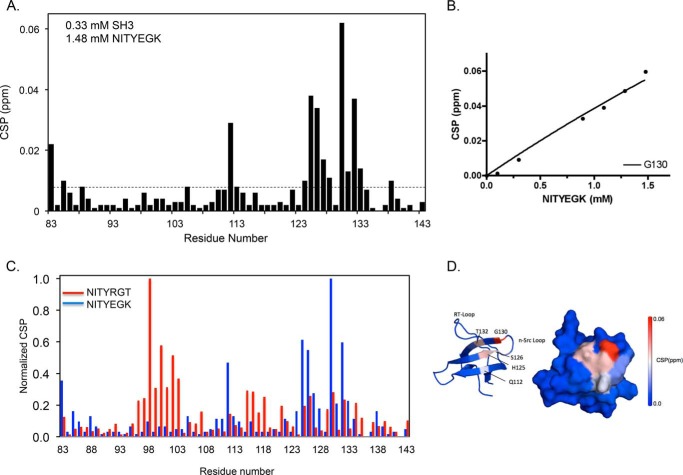
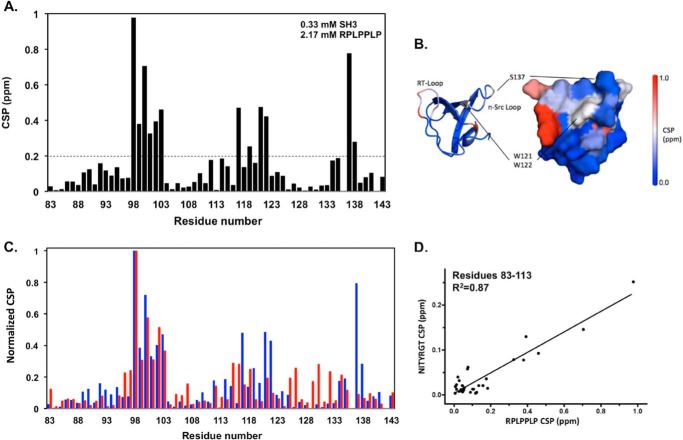
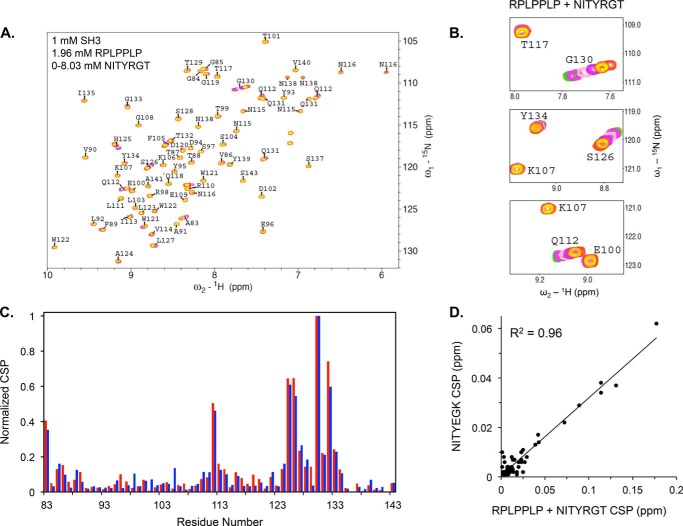
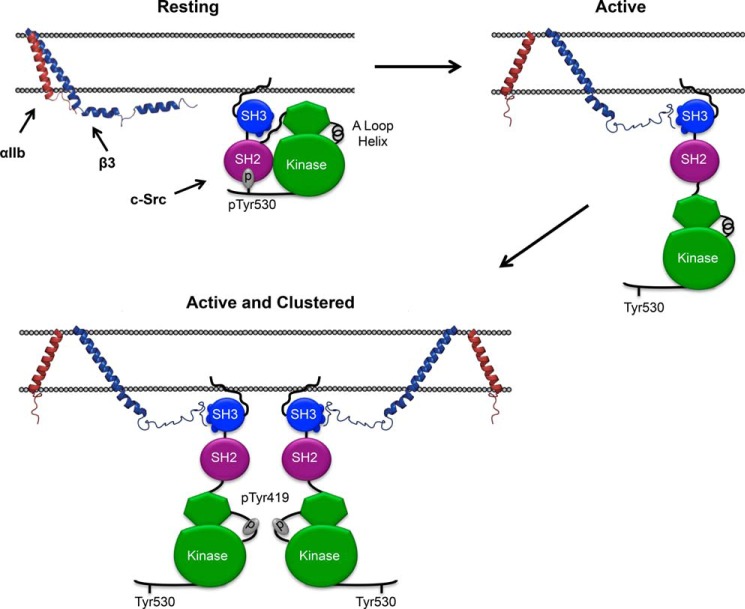
It is currently believed that inactive tyrosine kinase c-Src in platelets binds to the cytoplasmic tail of the β3 integrin subunit via its SH3 domain. Although a recent NMR study supports this contention, it is likely that such binding would be precluded in inactive c-Src because an auto-inhibitory linker physically occludes the β3 tail binding site. Accordingly, we have re-examined c-Src binding to β3 by immunoprecipitation as well as NMR spectroscopy. In unstimulated platelets, we detected little to no interaction between c-Src and β3. Following platelet activation, however, c-Src was co-immunoprecipitated with β3 in a time-dependent manner and underwent progressive activation as well. We then measured chemical shift perturbations in the (15)N-labeled SH3 domain induced by the C-terminal β3 tail peptide NITYRGT and found that the peptide interacted with the SH3 domain RT-loop and surrounding residues. A control peptide whose last three residues where replaced with those of the β1 cytoplasmic tail induced only small chemical shift perturbations on the opposite face of the SH3 domain. Next, to mimic inactive c-Src, we found that the canonical polyproline peptide RPLPPLP prevented binding of the β3 peptide to the RT- loop. Under these conditions, the β3 peptide induced chemical shift perturbations similar to the negative control. We conclude that the primary interaction of c-Src with the β3 tail occurs in its activated state and at a site that overlaps with PPII binding site in its SH3 domain. Interactions of inactive c-Src with β3 are weak and insensitive to β3 tail mutations.
Keywords: Src; Src homology 3 domain (SH3 domain); integrin; platelet; signal transduction.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
