

A novel insight into the cost-benefit model for the evolution of botanical carnivory
- PMID: 25948113
- PMCID: PMC4648460
- DOI: 10.1093/aob/mcv050
A novel insight into the cost-benefit model for the evolution of botanical carnivory
Abstract
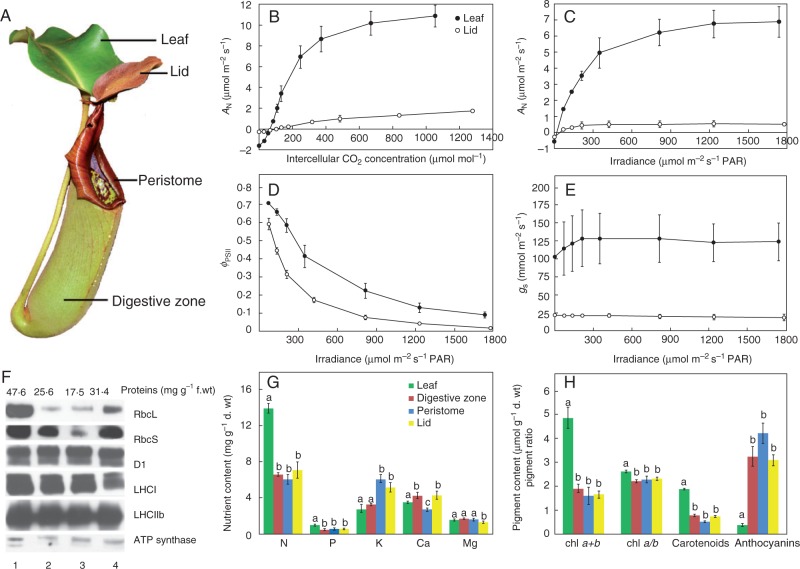
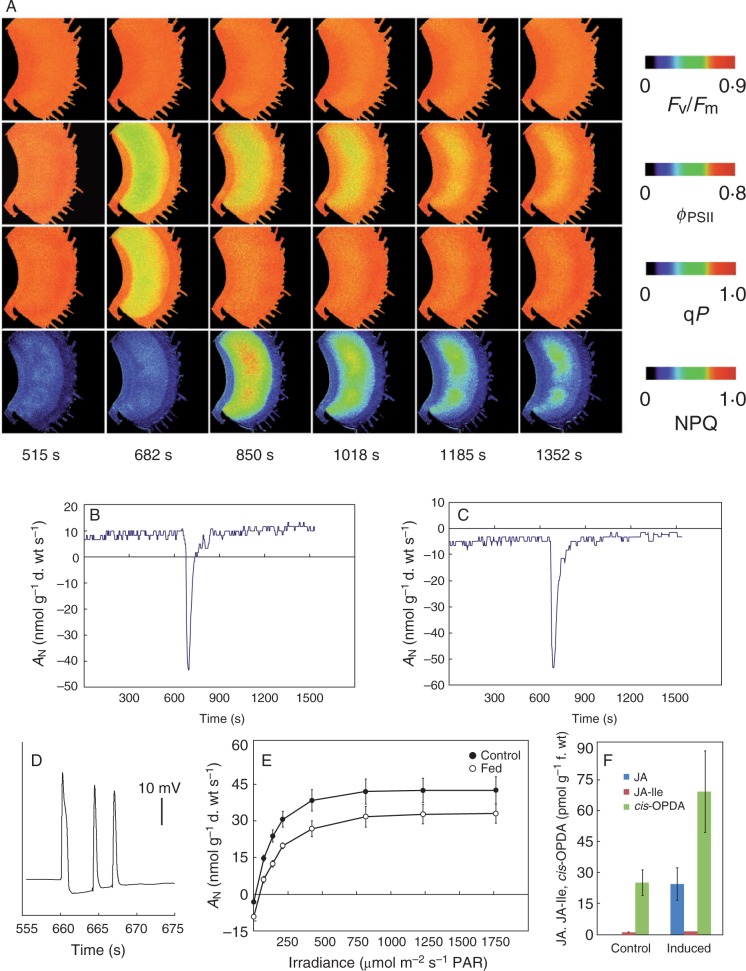
Background: The cost-benefit model for the evolution of botanical carnivory provides a conceptual framework for interpreting a wide range of comparative and experimental studies on carnivorous plants. This model assumes that the modified leaves called traps represent a significant cost for the plant, and this cost is outweighed by the benefits from increased nutrient uptake from prey, in terms of enhancing the rate of photosynthesis per unit leaf mass or area (AN) in the microsites inhabited by carnivorous plants.
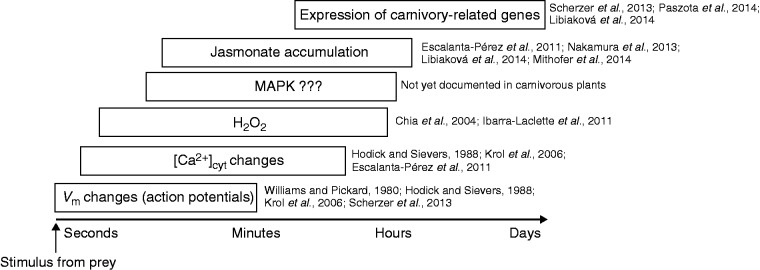
Scope: This review summarizes results from the classical interpretation of the cost-benefit model for evolution of botanical carnivory and highlights the costs and benefits of active trapping mechanisms, including water pumping, electrical signalling and accumulation of jasmonates. Novel alternative sequestration strategies (utilization of leaf litter and faeces) in carnivorous plants are also discussed in the context of the cost-benefit model.
Conclusions: Traps of carnivorous plants have lower AN than leaves, and the leaves have higher AN after feeding. Prey digestion, water pumping and electrical signalling represent a significant carbon cost (as an increased rate of respiration, RD) for carnivorous plants. On the other hand, jasmonate accumulation during the digestive period and reprogramming of gene expression from growth and photosynthesis to prey digestion optimizes enzyme production in comparison with constitutive secretion. This inducibility may have evolved as a cost-saving strategy beneficial for carnivorous plants. The similarities between plant defence mechanisms and botanical carnivory are highlighted.
Keywords: Action potential; Dionaea; Drosera; Nepenthes; Venus flytrap; botanical carnivory; carnivorous plant; cost–benefit; electrical signalling; jasmonates.
© The Author 2015. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures



References
-
- Adamec L. 1997. Mineral nutrition of carnivorous plants – a review. Botanical Review 63: 273–299.
-
- Adamec L. 2002. Leaf absorption of mineral nutrients in carnivorous plants stimulates root nutrient uptake. New Phytologist 155: 89–100. - PubMed
-
- Adamec L. 2003. Zero water flow in the carnivorous genus Genlisea. Carnivorous Plant Newsletter 32: 46–48.
-
- Adamec L. 2006. Respiration and photosynthesis of bladders and leaves of aquatic Utricularia species. Plant Biology 8: 765–769. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
