

Control of polarized assembly of actin filaments in cell motility
- PMID: 25948416
- PMCID: PMC4506460
- DOI: 10.1007/s00018-015-1914-2
Control of polarized assembly of actin filaments in cell motility
Abstract
Actin cytoskeleton remodeling, which drives changes in cell shape and motility, is orchestrated by a coordinated control of polarized assembly of actin filaments. Signal responsive, membrane-bound protein machineries initiate and regulate polarized growth of actin filaments by mediating transient links with their barbed ends, which elongate from polymerizable actin monomers. The barbed end of an actin filament thus stands out as a hotspot of regulation of filament assembly. It is the target of both soluble and membrane-bound agonists as well as antagonists of filament assembly. Here, we review the molecular mechanisms by which various regulators of actin dynamics bind, synergize or compete at filament barbed ends. Two proteins can compete for the barbed end via a mutually exclusive binding scheme. Alternatively, two regulators acting individually at barbed ends may be bound together transiently to terminal actin subunits at barbed ends, leading to the displacement of one by the other. The kinetics of these reactions is a key in understanding how filament length and membrane-filament linkage are controlled. It is also essential for understanding how force is produced to shape membranes by mechano-sensitive, processive barbed end tracking machineries like formins and by WASP-Arp2/3 branched filament arrays. A combination of biochemical and biophysical approaches, including bulk solution assembly measurements using pyrenyl-actin fluorescence, single filament dynamics, single molecule fluorescence imaging and reconstituted self-organized filament assemblies, have provided mechanistic insight into the role of actin polymerization in motile processes.
Figures
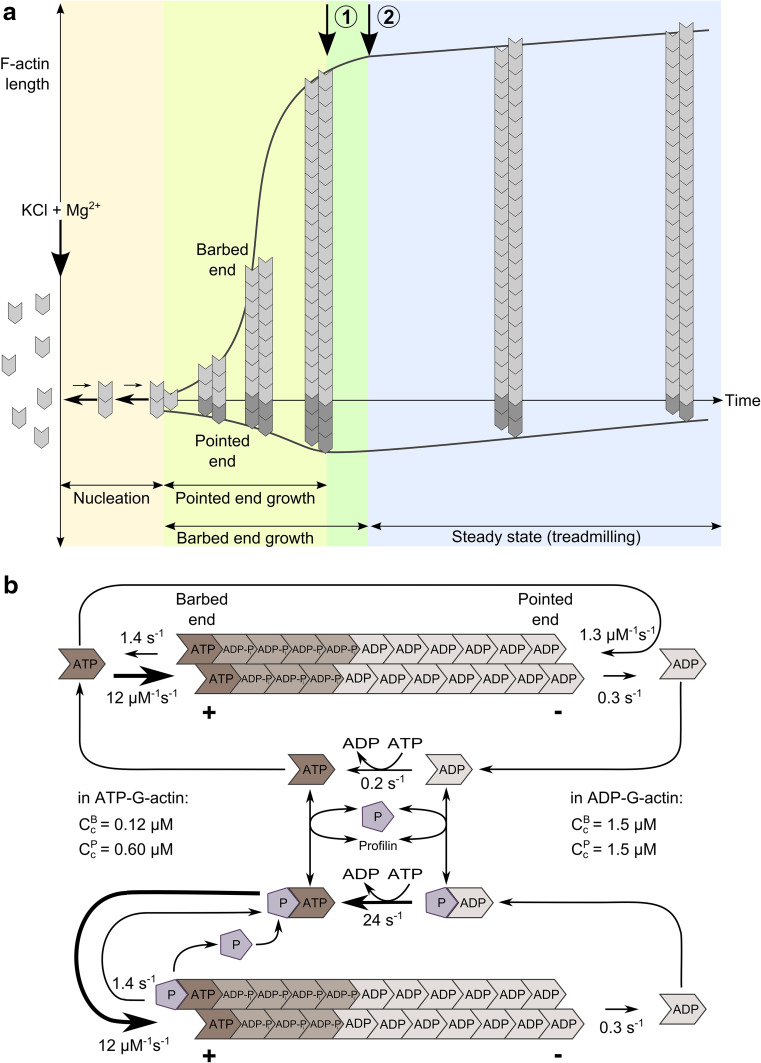
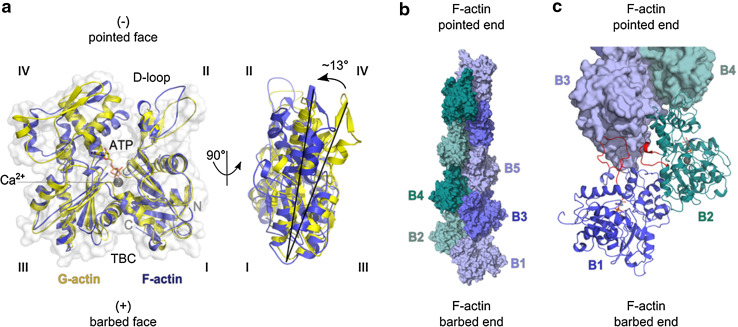
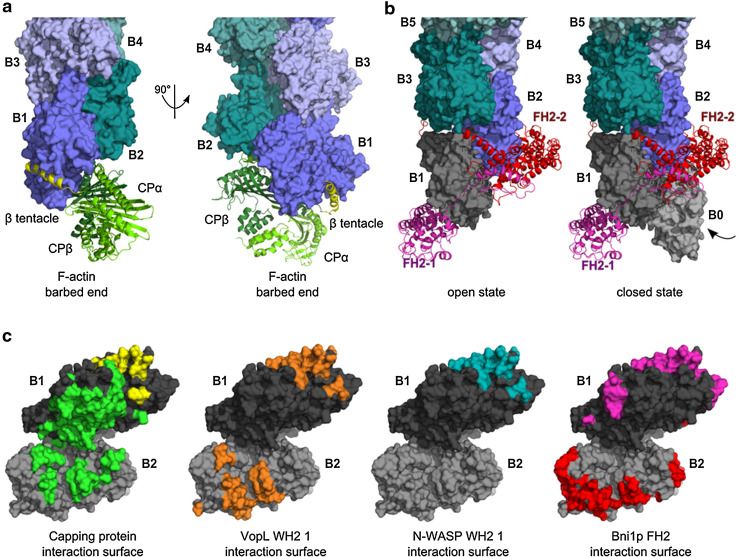
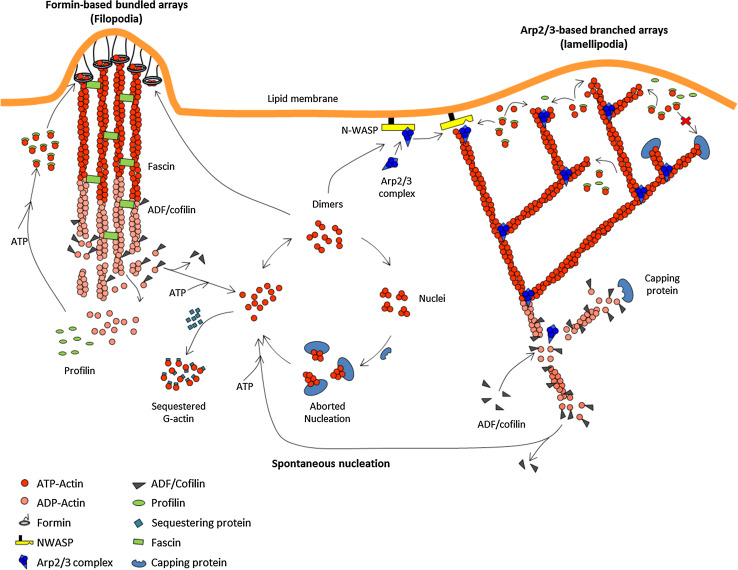
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
