

Natural Variation Identifies ICARUS1, a Universal Gene Required for Cell Proliferation and Growth at High Temperatures in Arabidopsis thaliana
- PMID: 25951176
- PMCID: PMC4423873
- DOI: 10.1371/journal.pgen.1005085
Natural Variation Identifies ICARUS1, a Universal Gene Required for Cell Proliferation and Growth at High Temperatures in Arabidopsis thaliana
Abstract
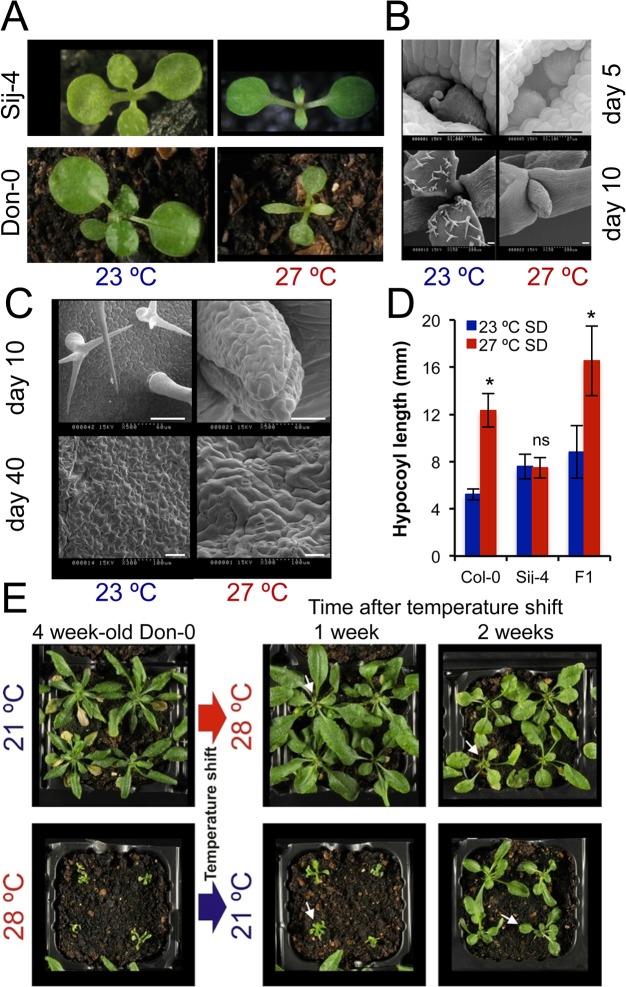
Plants are highly sensitive to environmental changes and even small variations in ambient temperature have severe consequences on their growth and development. Temperature affects multiple aspects of plant development, but the processes and mechanisms underlying thermo-sensitive growth responses are mostly unknown. Here we exploit natural variation in Arabidopsis thaliana to identify and characterize novel components and processes mediating thermo-sensitive growth responses in plants. Phenotypic screening of wild accessions identified several strains displaying pleiotropic growth defects, at cellular and organism levels, specifically at high ambient temperatures. Positional cloning and characterization of the underlying gene revealed that ICARUS1 (ICA1), which encodes a protein of the tRNAHis guanylyl transferase (Thg1) superfamily, is required for plant growth at high temperatures. Transcriptome and gene marker analyses together with DNA content measurements show that ICA1 loss-of-function results in down regulation of cell cycle associated genes at high temperatures, which is linked with a block in G2/M transition and endoreduplication. In addition, plants with mutations in ICA1 show enhanced sensitivity to DNA damage. Characterization of additional strains that carry lesions in ICA1, but display normal growth, shows that alternative splicing is likely to alleviate the deleterious effects of some natural mutations. Furthermore, analyses of worldwide and regional collections of natural accessions indicate that ICA1 loss-of-function has arisen several times independently, and that these occur at high frequency in some local populations. Overall our results suggest that ICA1-mediated-modulation of fundamental processes such as tRNAHis maturation, modify plant growth responses to temperature changes in a quantitative and reversible manner, in natural populations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Genetic Architecture of Natural Variation in Thermal Responses of Arabidopsis.Plant Physiol. 2015 Sep;169(1):647-59. doi: 10.1104/pp.15.00942. Epub 2015 Jul 20. Plant Physiol. 2015. PMID: 26195568 Free PMC article.
-
Genetic Interactions and Molecular Evolution of the Duplicated Genes ICARUS2 and ICARUS1 Help Arabidopsis Plants Adapt to Different Ambient Temperatures.Plant Cell. 2019 Jun;31(6):1222-1237. doi: 10.1105/tpc.18.00938. Epub 2019 Apr 16. Plant Cell. 2019. PMID: 30992321 Free PMC article.
-
ASYMMETRIC LEAVES1, an Arabidopsis gene that is involved in the control of cell differentiation in leaves.Planta. 2002 Mar;214(5):694-702. doi: 10.1007/s004250100673. Epub 2001 Nov 10. Planta. 2002. PMID: 11882937
-
[Arabidopsis thaliana accessions - a tool for biochemical and phylogentical studies].Postepy Biochem. 2015;61(1):102-13. Postepy Biochem. 2015. PMID: 26281359 Review. Polish.
-
Exploiting natural variation to uncover candidate genes that control element accumulation in Arabidopsis thaliana.New Phytol. 2012 Mar;193(4):859-66. doi: 10.1111/j.1469-8137.2011.03977.x. New Phytol. 2012. PMID: 22403822 Review.
Cited by
-
Evolution of Plant Phenotypes, from Genomes to Traits.G3 (Bethesda). 2016 Apr;6(4):775-778. doi: 10.1534/g3.115.025502. Epub 2016 Feb 11. G3 (Bethesda). 2016. PMID: 26869620 Free PMC article. No abstract available.
-
Genetic Architecture of Natural Variation in Thermal Responses of Arabidopsis.Plant Physiol. 2015 Sep;169(1):647-59. doi: 10.1104/pp.15.00942. Epub 2015 Jul 20. Plant Physiol. 2015. PMID: 26195568 Free PMC article.
-
Dynamic mRNP Remodeling in Response to Internal and External Stimuli.Biomolecules. 2020 Sep 11;10(9):1310. doi: 10.3390/biom10091310. Biomolecules. 2020. PMID: 32932892 Free PMC article. Review.
-
High temperature restricts cell division and leaf size by coordination of PIF4 and TCP4 transcription factors.Plant Physiol. 2022 Nov 28;190(4):2380-2397. doi: 10.1093/plphys/kiac345. Plant Physiol. 2022. PMID: 35880840 Free PMC article.
-
Template-dependent nucleotide addition in the reverse (3'-5') direction by Thg1-like protein.Sci Adv. 2016 Mar 25;2(3):e1501397. doi: 10.1126/sciadv.1501397. eCollection 2016 Mar. Sci Adv. 2016. PMID: 27051866 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
