

Neuronal control of locomotor handedness in Drosophila
- PMID: 25953337
- PMCID: PMC4450378
- DOI: 10.1073/pnas.1500804112
Neuronal control of locomotor handedness in Drosophila
Abstract
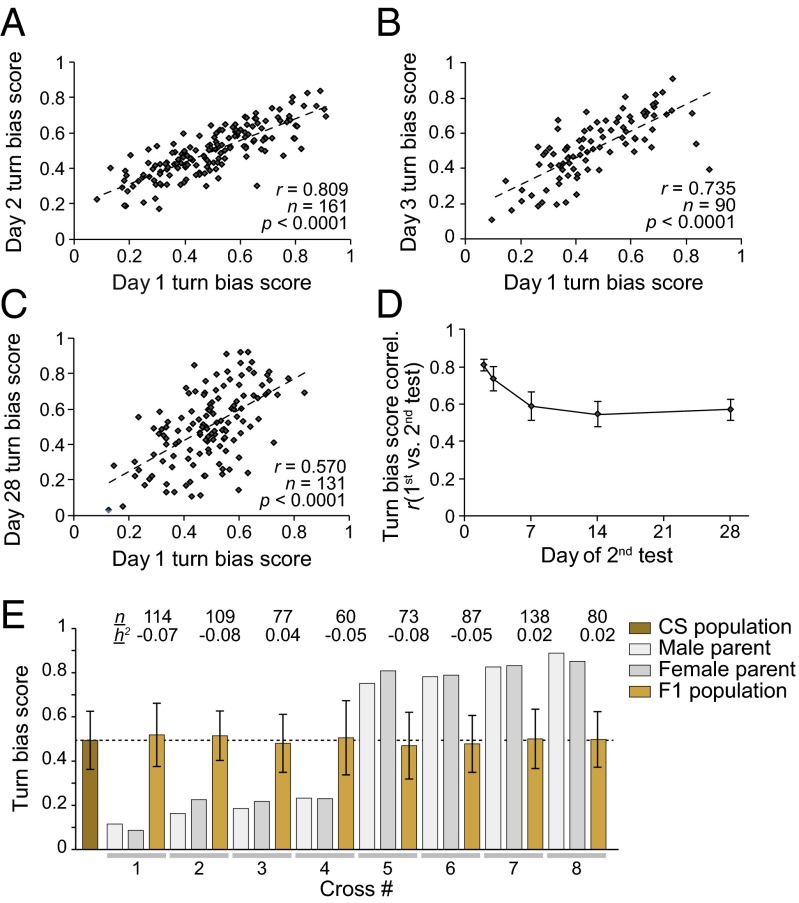
Genetically identical individuals display variability in their physiology, morphology, and behaviors, even when reared in essentially identical environments, but there is little mechanistic understanding of the basis of such variation. Here, we investigated whether Drosophila melanogaster displays individual-to-individual variation in locomotor behaviors. We developed a new high-throughout platform capable of measuring the exploratory behavior of hundreds of individual flies simultaneously. With this approach, we find that, during exploratory walking, individual flies exhibit significant bias in their left vs. right locomotor choices, with some flies being strongly left biased or right biased. This idiosyncrasy was present in all genotypes examined, including wild-derived populations and inbred isogenic laboratory strains. The biases of individual flies persist for their lifetime and are nonheritable: i.e., mating two left-biased individuals does not yield left-biased progeny. This locomotor handedness is uncorrelated with other asymmetries, such as the handedness of gut twisting, leg-length asymmetry, and wing-folding preference. Using transgenics and mutants, we find that the magnitude of locomotor handedness is under the control of columnar neurons within the central complex, a brain region implicated in motor planning and execution. When these neurons are silenced, exploratory laterality increases, with more extreme leftiness and rightiness. This observation intriguingly implies that the brain may be able to dynamically regulate behavioral individuality.
Keywords: behavior; central complex; circuit mapping; individuality; personality.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Souman JL, Frissen I, Sreenivasa MN, Ernst MO. Walking straight into circles. Curr Biol. 2009;19(18):1538–1542. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
