

Alternative activation of macrophages and pulmonary fibrosis are modulated by scavenger receptor, macrophage receptor with collagenous structure
- PMID: 25953850
- PMCID: PMC4511206
- DOI: 10.1096/fj.15-271304
Alternative activation of macrophages and pulmonary fibrosis are modulated by scavenger receptor, macrophage receptor with collagenous structure
Abstract
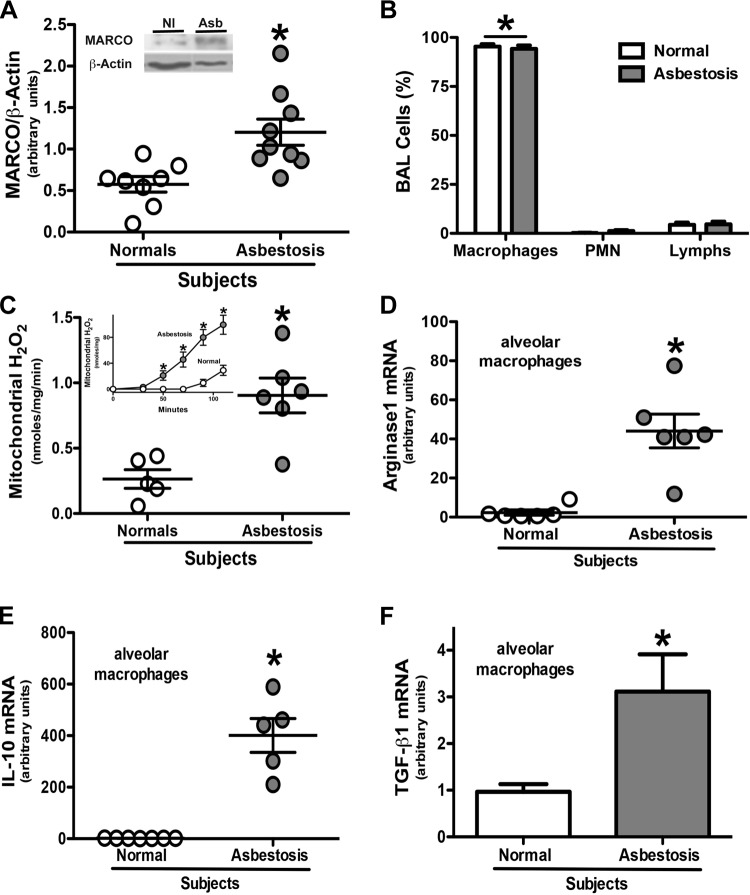
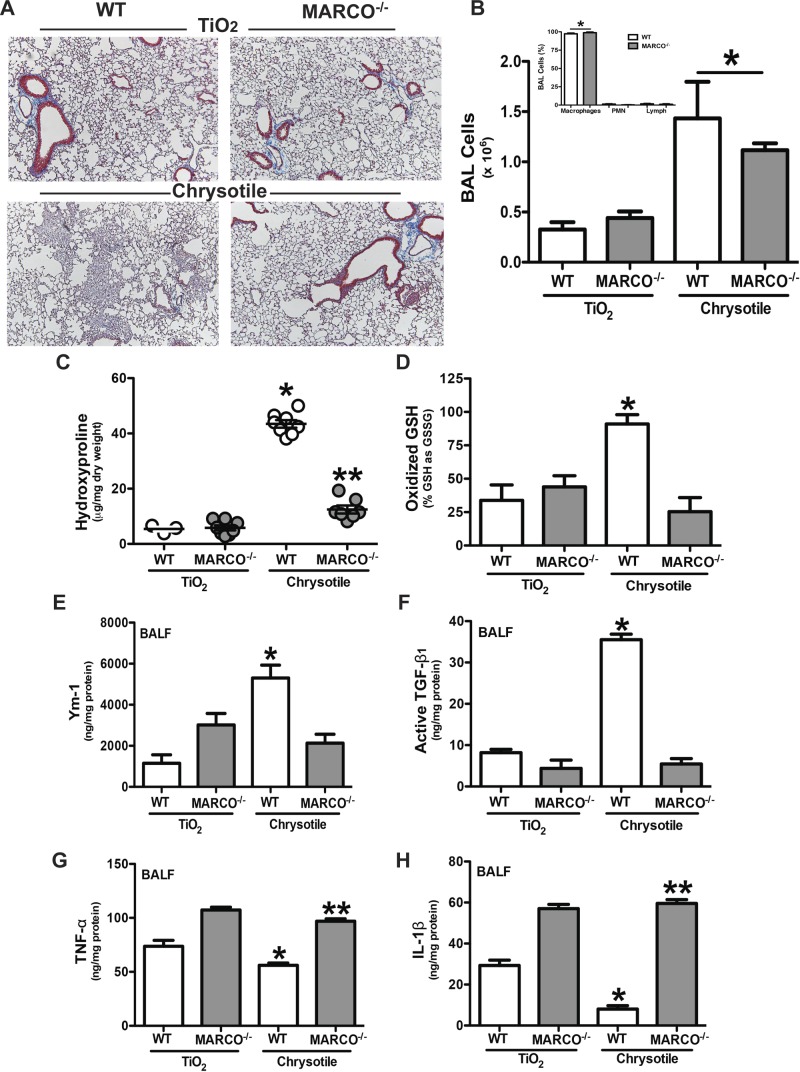
Alternative activation of alveolar macrophages is linked to fibrosis following exposure to asbestos. The scavenger receptor, macrophage receptor with collagenous structure (MARCO), provides innate immune defense against inhaled particles and pathogens; however, a receptor for asbestos has not been identified. We hypothesized that MARCO acts as an initial signaling receptor for asbestos, polarizes macrophages to a profibrotic M2 phenotype, and is required for the development of asbestos-induced fibrosis. Compared with normal subjects, alveolar macrophages isolated from patients with asbestosis express higher amounts of MARCO and have greater profibrotic polarization. Arginase 1 (40-fold) and IL-10 (265-fold) were higher in patients. In vivo, the genetic deletion of MARCO attenuated the profibrotic environment and pulmonary fibrosis in mice exposed to chrysotile. Moreover, alveolar macrophages from MARCO(-/-) mice polarize to an M1 phenotype, whereas wild-type mice have higher Ym1 (>3.0-fold) and nearly 7-fold more active TGF-β1 in bronchoalveolar lavage (BAL) fluid (BALF). Arg(432) and Arg(434) in domain V of MARCO are required for the polarization of macrophages to a profibrotic phenotype as mutation of these residues reduced FIZZ1 expression (17-fold) compared with cells expressing MARCO. These observations demonstrate that a macrophage membrane protein regulates the fibrotic response to lung injury and suggest a novel target for therapeutic intervention.
Keywords: MARCO; asbestos; mitochondria.
© FASEB.
Figures



References
-
- Gordon S., Martinez F. O. (2010) Alternative activation of macrophages: mechanism and functions. Immunity 32, 593–604 - PubMed
-
- Martinez F. O., Helming L., Gordon S. (2009) Alternative activation of macrophages: an immunologic functional perspective. Annu. Rev. Immunol. 27, 451–483 - PubMed
-
- Herbert D. R., Hölscher C., Mohrs M., Arendse B., Schwegmann A., Radwanska M., Leeto M., Kirsch R., Hall P., Mossmann H., Claussen B., Förster I., Brombacher F. (2004) Alternative macrophage activation is essential for survival during schistosomiasis and downmodulates T helper 1 responses and immunopathology. Immunity 20, 623–635 - PubMed
-
- Nair M. G., Du Y., Perrigoue J. G., Zaph C., Taylor J. J., Goldschmidt M., Swain G. P., Yancopoulos G. D., Valenzuela D. M., Murphy A., Karow M., Stevens S., Pearce E. J., Artis D. (2009) Alternatively activated macrophage-derived RELM-alpha is a negative regulator of type 2 inflammation in the lung. J. Exp. Med. 206, 937–952 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
