

The functional basis of wing patterning in Heliconius butterflies: the molecules behind mimicry
- PMID: 25953905
- PMCID: PMC4423356
- DOI: 10.1534/genetics.114.172387
The functional basis of wing patterning in Heliconius butterflies: the molecules behind mimicry
Abstract
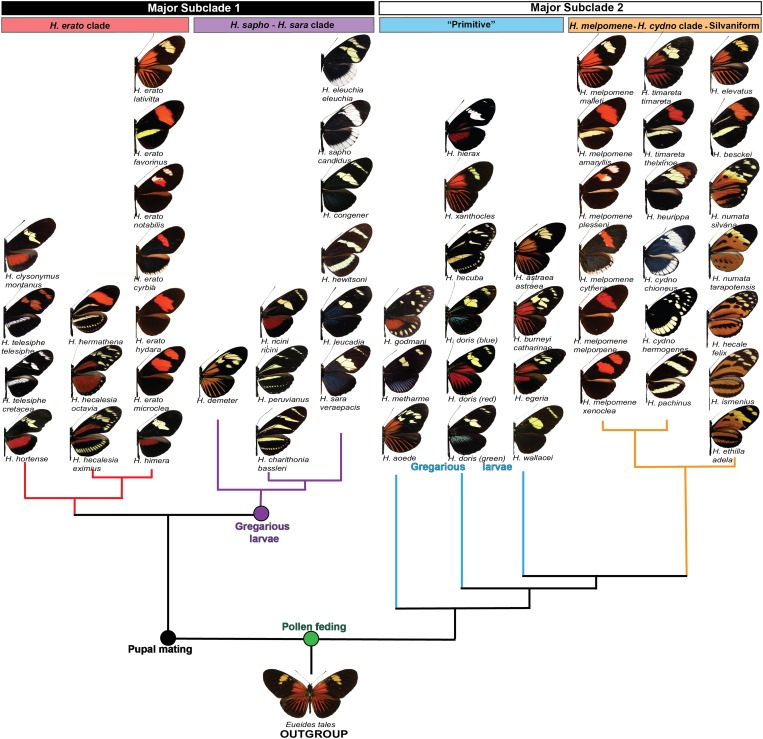
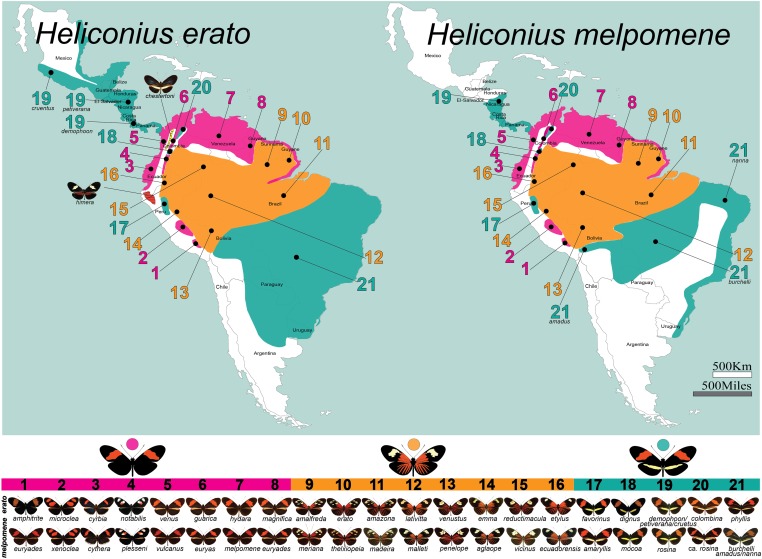
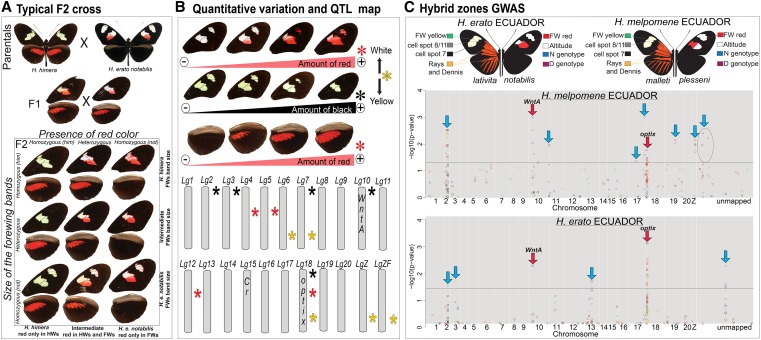
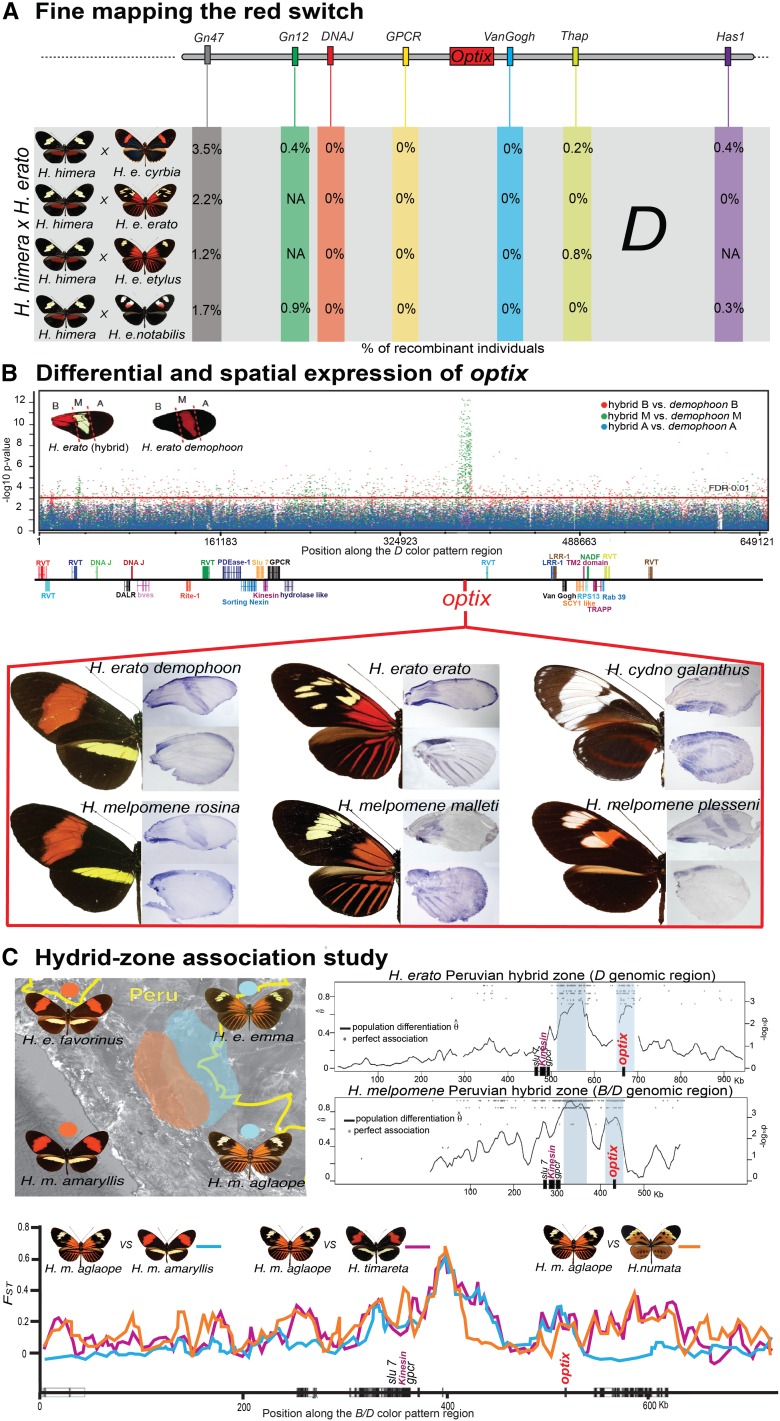
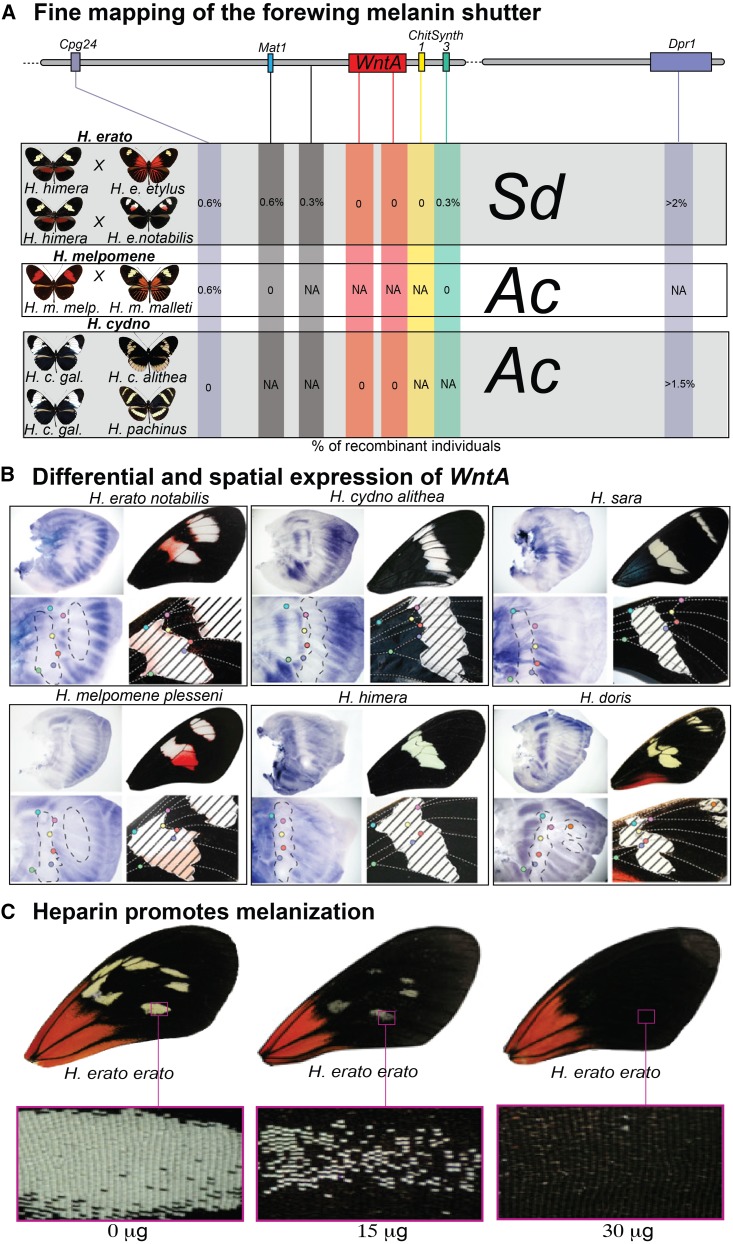
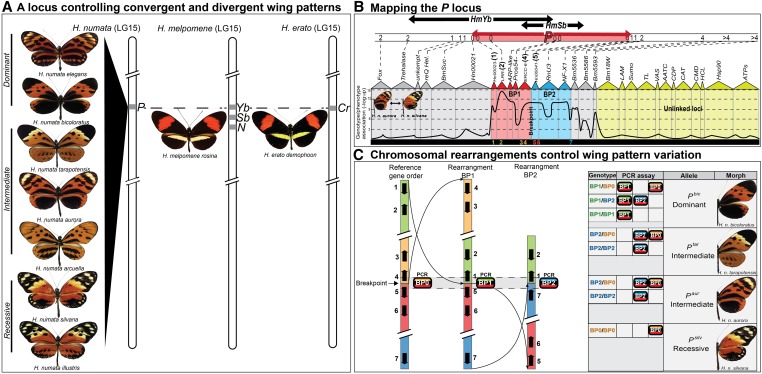
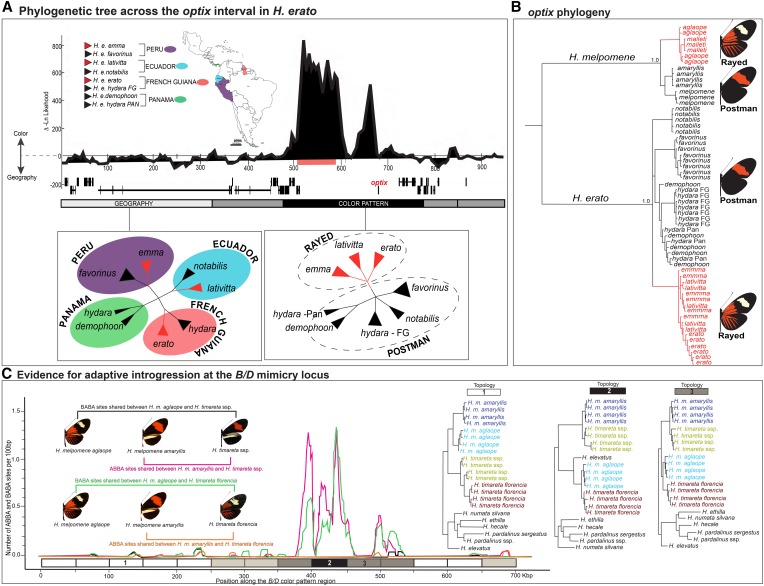
Wing-pattern mimicry in butterflies has provided an important example of adaptation since Charles Darwin and Alfred Russell Wallace proposed evolution by natural selection >150 years ago. The neotropical butterfly genus Heliconius played a central role in the development of mimicry theory and has since been studied extensively in the context of ecology and population biology, behavior, and mimicry genetics. Heliconius species are notable for their diverse color patterns, and previous crossing experiments revealed that much of this variation is controlled by a small number of large-effect, Mendelian switch loci. Recent comparative analyses have shown that the same switch loci control wing-pattern diversity throughout the genus, and a number of these have now been positionally cloned. Using a combination of comparative genetic mapping, association tests, and gene expression analyses, variation in red wing patterning throughout Heliconius has been traced back to the action of the transcription factor optix. Similarly, the signaling ligand WntA has been shown to control variation in melanin patterning across Heliconius and other butterflies. Our understanding of the molecular basis of Heliconius mimicry is now providing important insights into a variety of additional evolutionary phenomena, including the origin of supergenes, the interplay between constraint and evolvability, the genetic basis of convergence, the potential for introgression to facilitate adaptation, the mechanisms of hybrid speciation in animals, and the process of ecological speciation.
Keywords: Heliconius; adaptation; mimicry; speciation.
Copyright © 2015 by the Genetics Society of America.
Figures
References
-
- Bates H. W., 1862. Contributions to an insect fauna of the Amazon Valley. Lepidoptera: Heliconidae. Trans. Linn. Soc. Lond. 23: 495–566.
-
- Bates H. W., 1864. The Naturalist on the River Amazons. A Record of Adventures, Habits of Animals, Sketches of Brazilian and Indian life, and Aspects of Nature Under the Equator, During Eleven Years of Travel. J. Murray, London.
-
- Baxter S. W., Johnston S. E., Jiggins C. D., 2009. Butterfly speciation and the distribution of gene effect sizes fixed during adaptation. Heredity 102: 57–65. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
