

Shape representations in the primate dorsal visual stream
- PMID: 25954189
- PMCID: PMC4406065
- DOI: 10.3389/fncom.2015.00043
Shape representations in the primate dorsal visual stream
Abstract
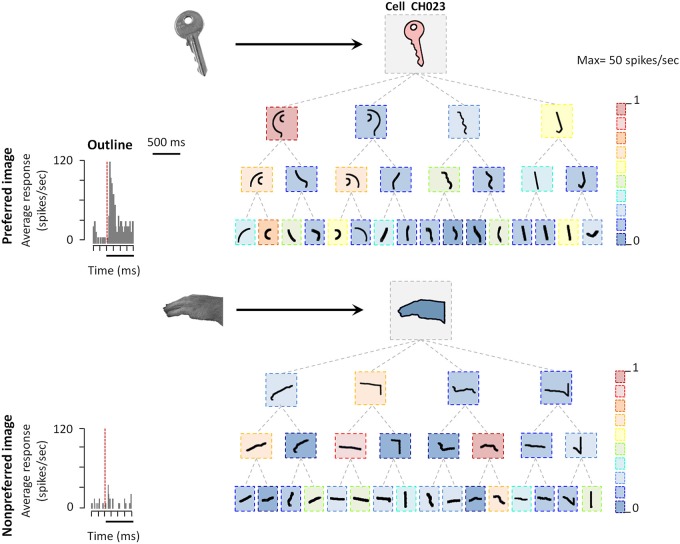
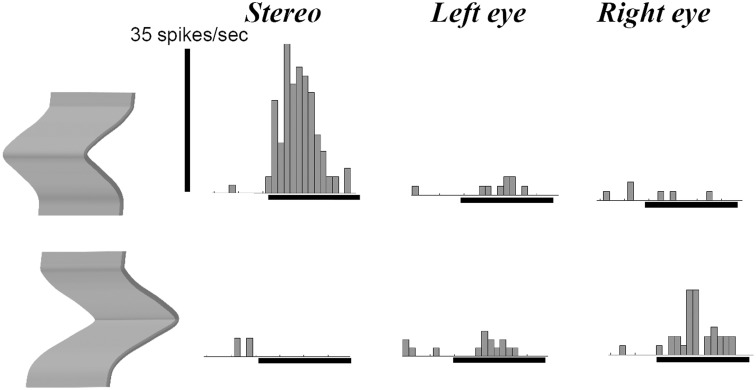
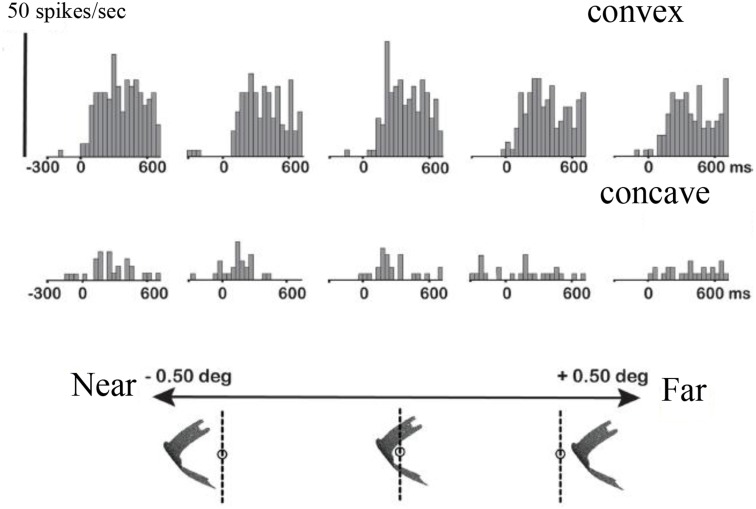
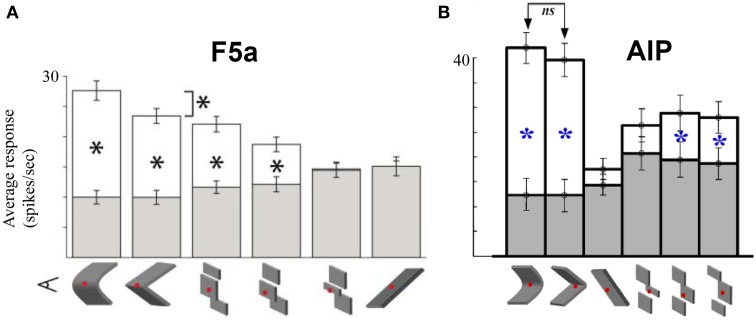
The primate visual system extracts object shape information for object recognition in the ventral visual stream. Recent research has demonstrated that object shape is also processed in the dorsal visual stream, which is specialized for spatial vision and the planning of actions. A number of studies have investigated the coding of 2D shape in the anterior intraparietal area (AIP), one of the end-stage areas of the dorsal stream which has been implicated in the extraction of affordances for the purpose of grasping. These findings challenge the current understanding of area AIP as a critical stage in the dorsal stream for the extraction of object affordances. The representation of three-dimensional (3D) shape has been studied in two interconnected areas known to be critical for object grasping: area AIP and area F5a in the ventral premotor cortex (PMv), to which AIP projects. In both areas neurons respond selectively to 3D shape defined by binocular disparity, but the latency of the neural selectivity is approximately 10 ms longer in F5a compared to AIP, consistent with its higher position in the hierarchy of cortical areas. Furthermore, F5a neurons were more sensitive to small amplitudes of 3D curvature and could detect subtle differences in 3D structure more reliably than AIP neurons. In both areas, 3D-shape selective neurons were co-localized with neurons showing motor-related activity during object grasping in the dark, indicating a close convergence of visual and motor information on the same clusters of neurons.
Keywords: depth; dorsal stream; macaque; object; parietal cortex; shape; visual cortex.
Figures


References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous