

Making sense of transcription networks
- PMID: 25957680
- PMCID: PMC4531093
- DOI: 10.1016/j.cell.2015.04.014
Making sense of transcription networks
Abstract
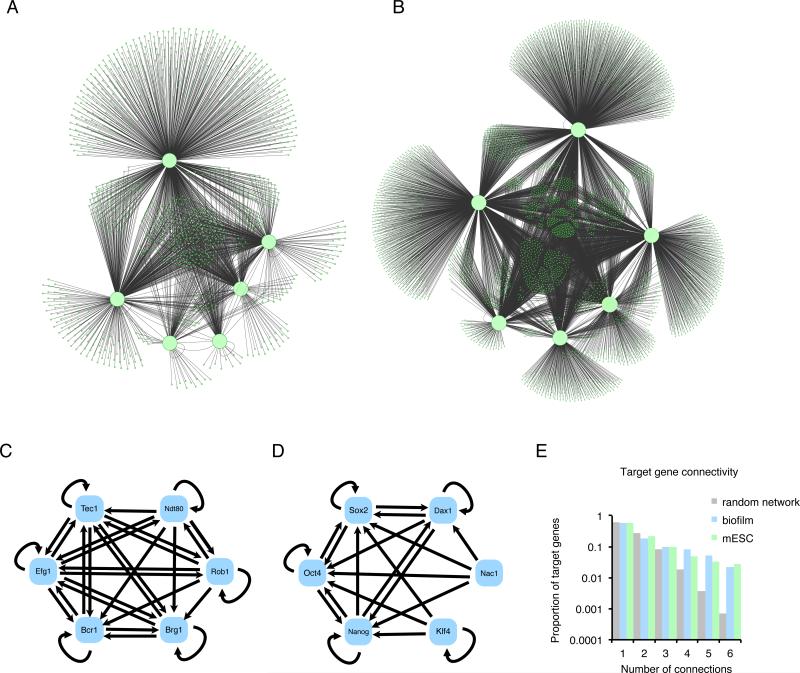
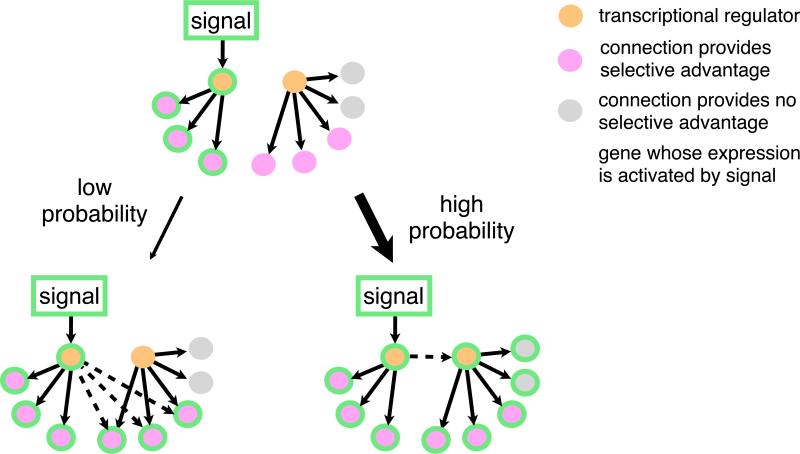
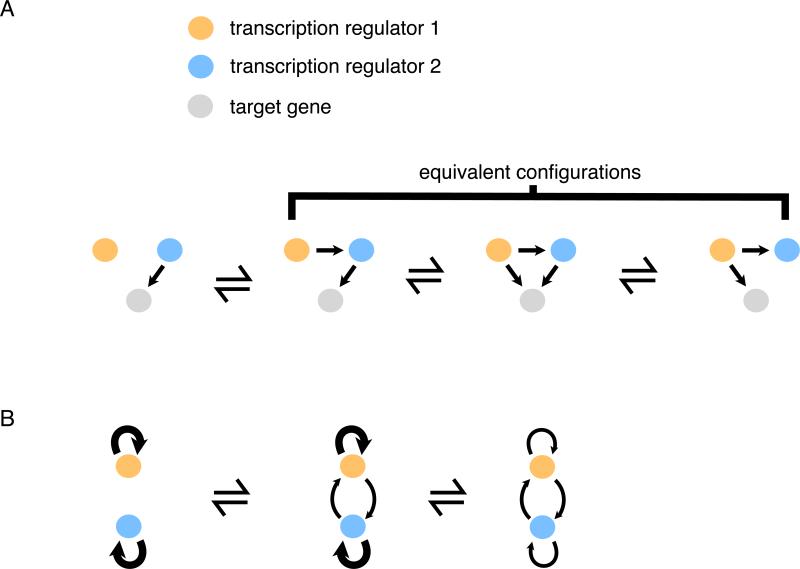
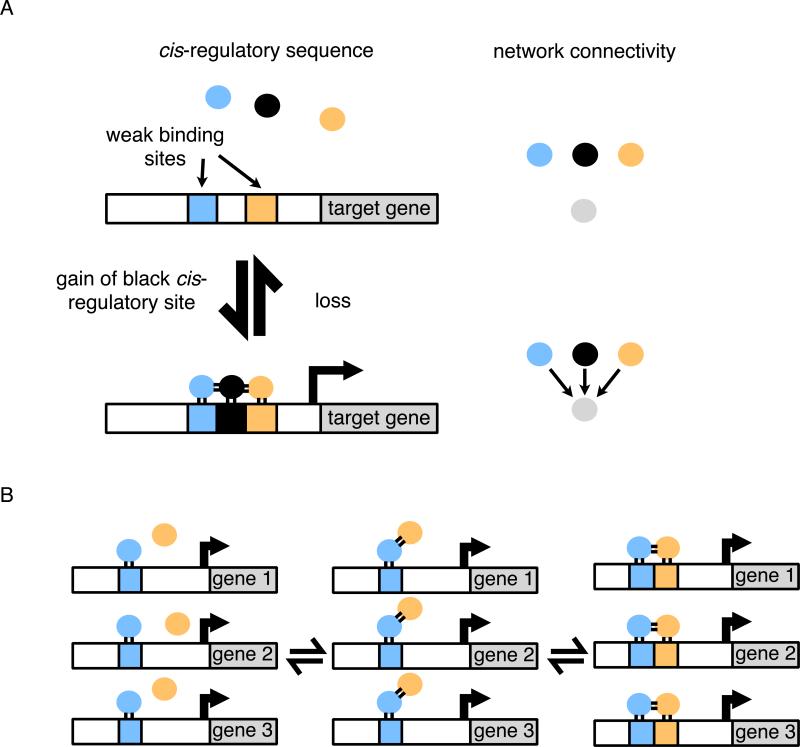
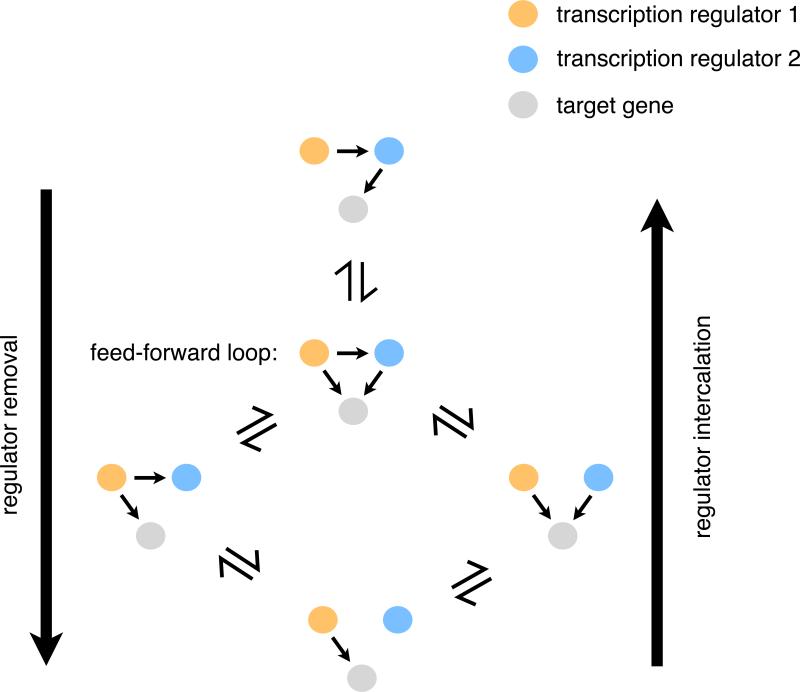
When transcription regulatory networks are compared among distantly related eukaryotes, a number of striking similarities are observed: a larger-than-expected number of genes, extensive overlapping connections, and an apparently high degree of functional redundancy. It is often assumed that the complexity of these networks represents optimized solutions, precisely sculpted by natural selection; their common features are often asserted to be adaptive. Here, we discuss support for an alternative hypothesis: the common structural features of transcription networks arise from evolutionary trajectories of "least resistance"--that is, the relative ease with which certain types of network structures are formed during their evolution.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
