

Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement
- PMID: 25957686
- PMCID: PMC4457382
- DOI: 10.1016/j.cell.2015.02.053
Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement
Abstract
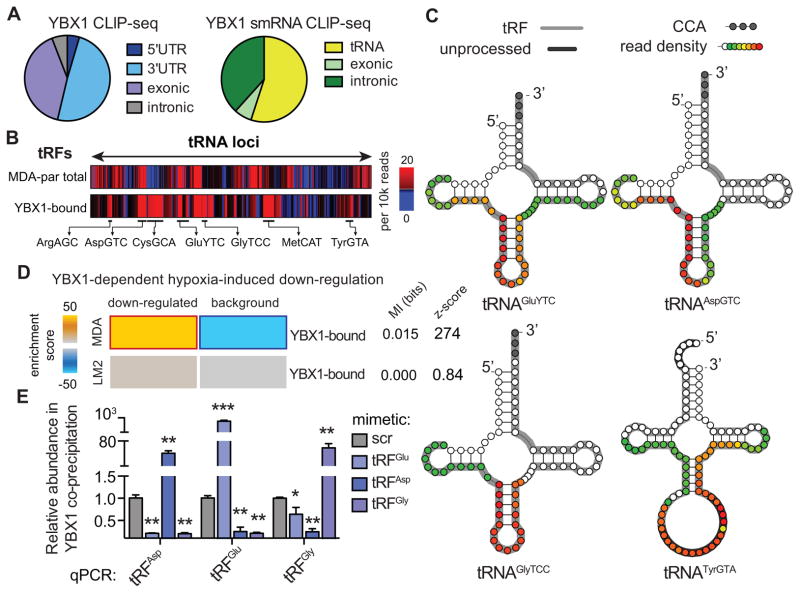
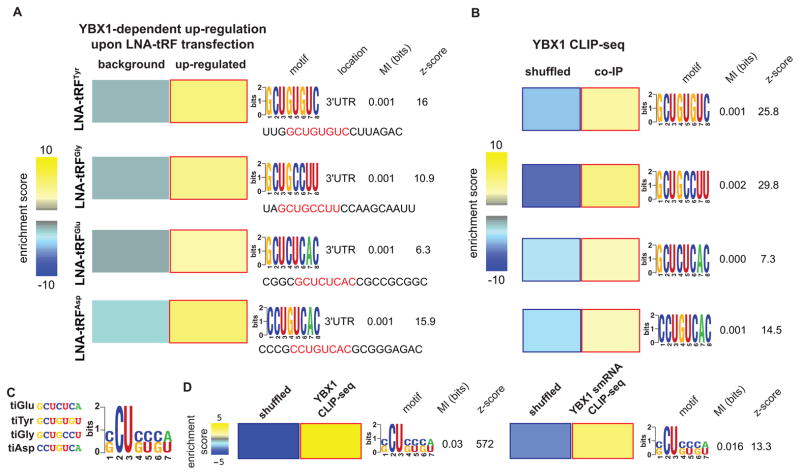
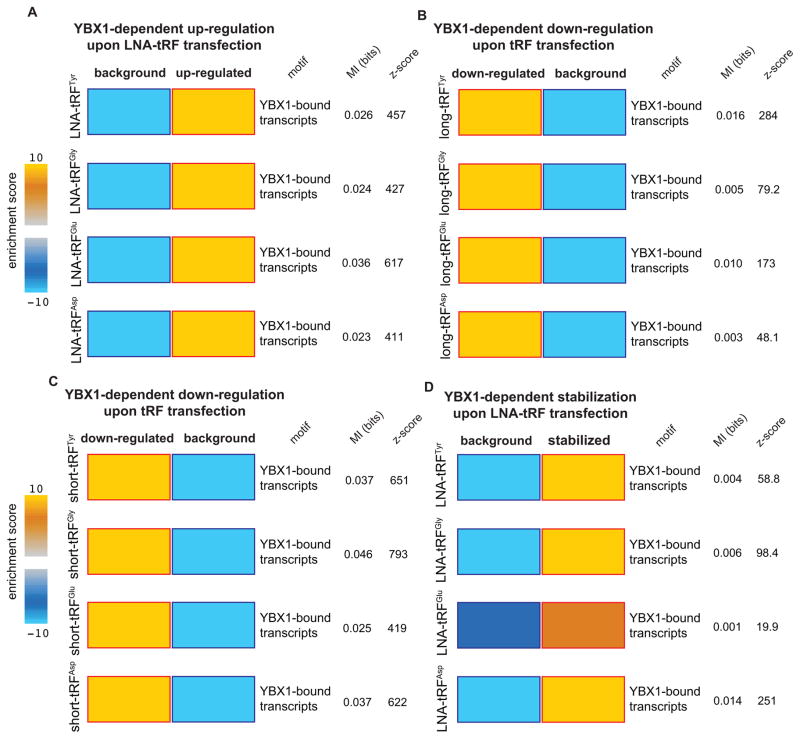
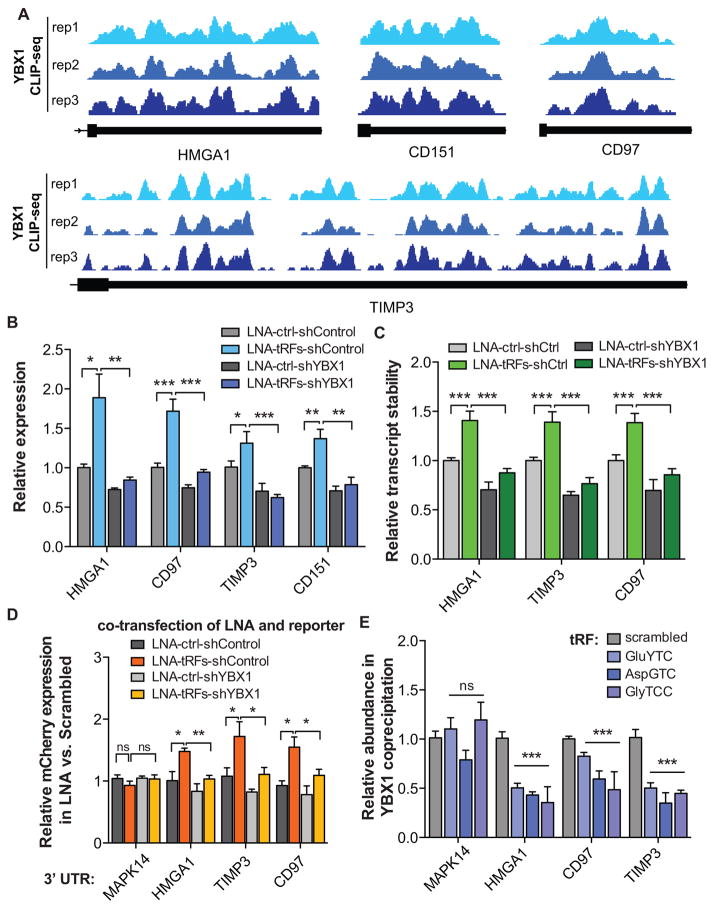
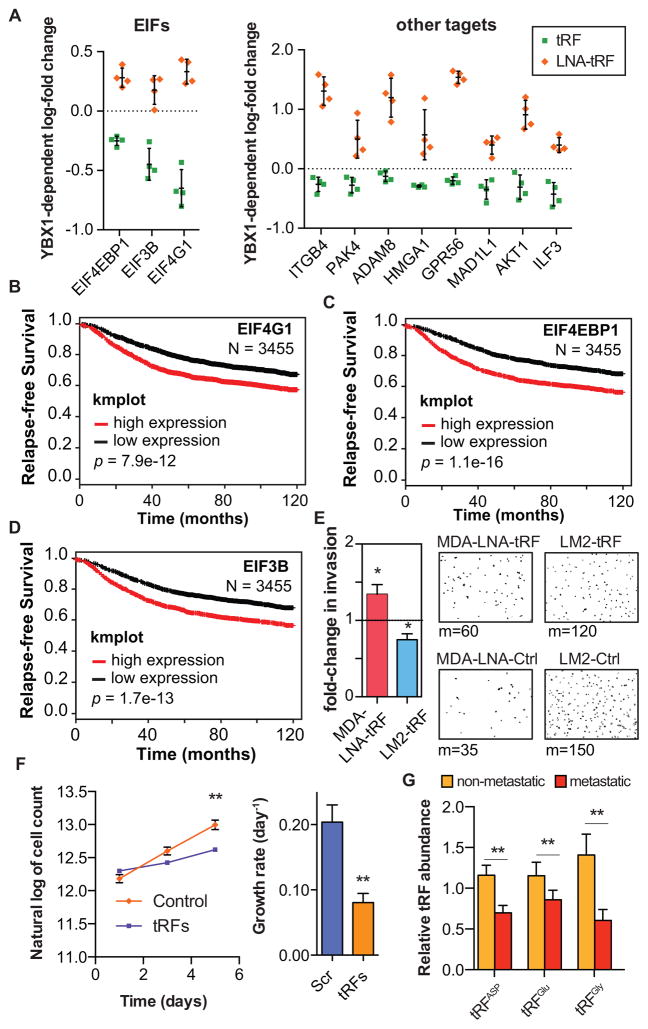
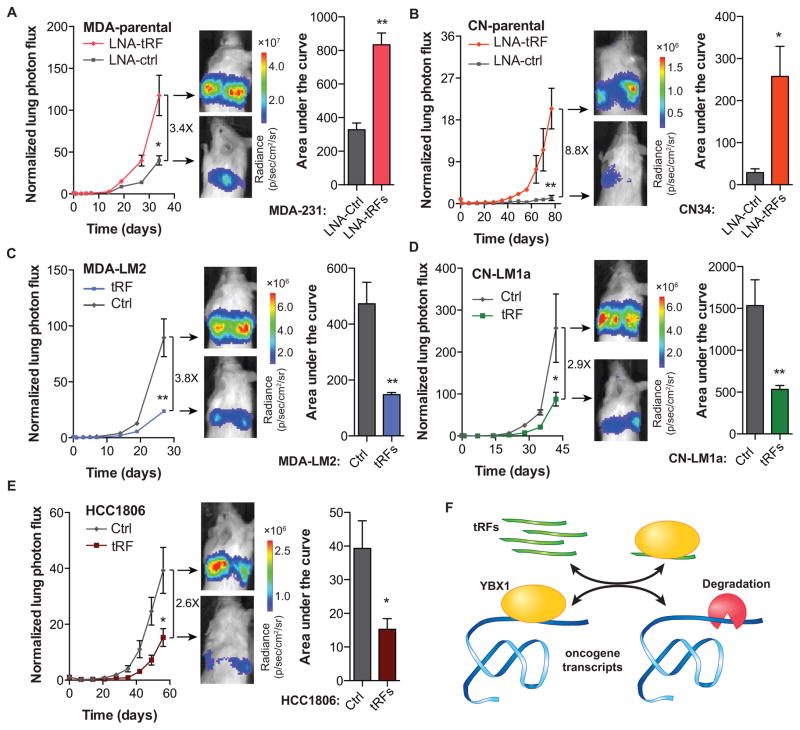
Upon exposure to stress, tRNAs are enzymatically cleaved, yielding distinct classes of tRNA-derived fragments (tRFs), yielding distinct classes of tRFs. We identify a novel class of tRFs derived from tRNA(Glu), tRNA(Asp), tRNA(Gly), and tRNA(Tyr) that, upon induction, suppress the stability of multiple oncogenic transcripts in breast cancer cells by displacing their 3' untranslated regions (UTRs) from the RNA-binding protein YBX1. This mode of post-transcriptional silencing is sequence specific, as these fragments all share a common motif that matches the YBX1 recognition sequence. Loss-of-function and gain-of-function studies, using anti-sense locked-nucleic acids (LNAs) and synthetic RNA mimetics, respectively, revealed that these fragments suppress growth under serum-starvation, cancer cell invasion, and metastasis by breast cancer cells. Highly metastatic cells evade this tumor-suppressive pathway by attenuating the induction of these tRFs. Our findings reveal a tumor-suppressive role for specific tRNA-derived fragments and describe a molecular mechanism for their action. This transcript displacement-based mechanism may generalize to other tRNA, ribosomal-RNA, and sno-RNA fragments.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures

Comment in
-
Non-coding RNA: Stressed to bits.Nat Rev Cancer. 2015 Jun;15(6):320. doi: 10.1038/nrc3966. Nat Rev Cancer. 2015. PMID: 25998710 No abstract available.
References
-
- Borek E, Baliga BS, Gehrke CW, Kuo CW, Belman S, Troll W, Waalkes TP. High turnover rate of transfer RNA in tumor tissue. Cancer research. 1977;37:3362–3366. - PubMed
-
- Bristow RG, Hill RP. Hypoxia and metabolism. Hypoxia, DNA repair and genetic instability. Nature reviews Cancer. 2008;8:180–192. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
