

Structural snapshots of actively translating human ribosomes
- PMID: 25957688
- PMCID: PMC4432480
- DOI: 10.1016/j.cell.2015.03.052
Structural snapshots of actively translating human ribosomes
Abstract
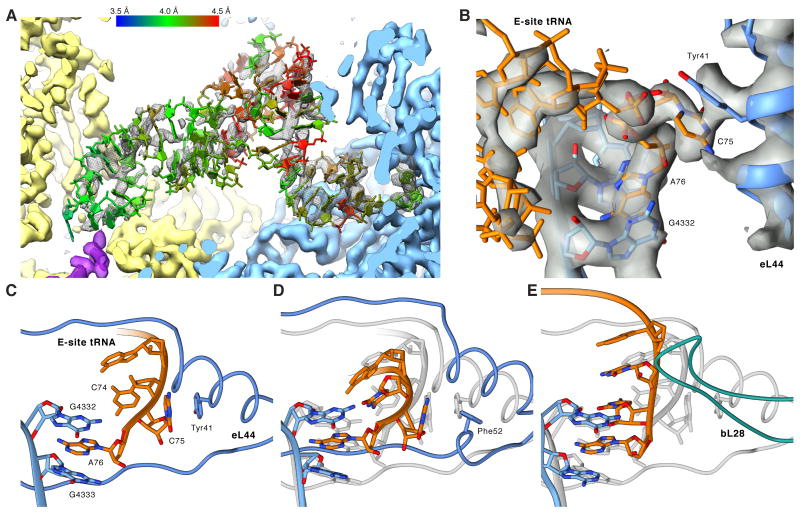
Macromolecular machines, such as the ribosome, undergo large-scale conformational changes during their functional cycles. Although their mode of action is often compared to that of mechanical machines, a crucial difference is that, at the molecular dimension, thermodynamic effects dominate functional cycles, with proteins fluctuating stochastically between functional states defined by energetic minima on an energy landscape. Here, we have used cryo-electron microscopy to image ex-vivo-derived human polysomes as a source of actively translating ribosomes. Multiparticle refinement and 3D variability analysis allowed us to visualize a variety of native translation intermediates. Significantly populated states include not only elongation cycle intermediates in pre- and post-translocational states, but also eEF1A-containing decoding and termination/recycling complexes. Focusing on the post-translocational state, we extended this assessment to the single-residue level, uncovering striking details of ribosome-ligand interactions and identifying both static and functionally important dynamic elements.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures

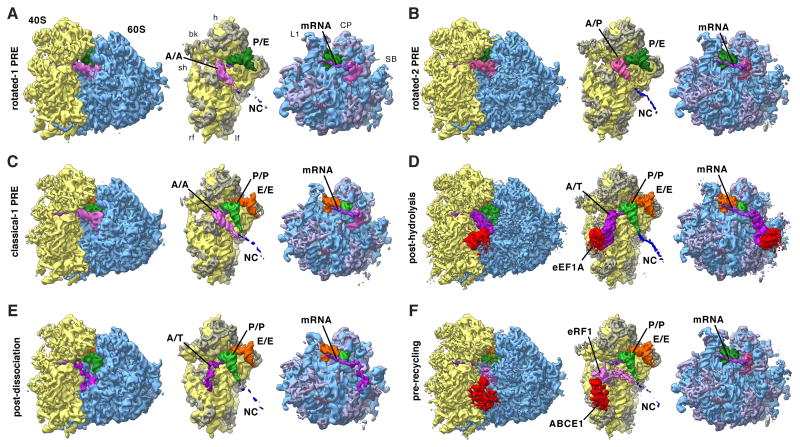
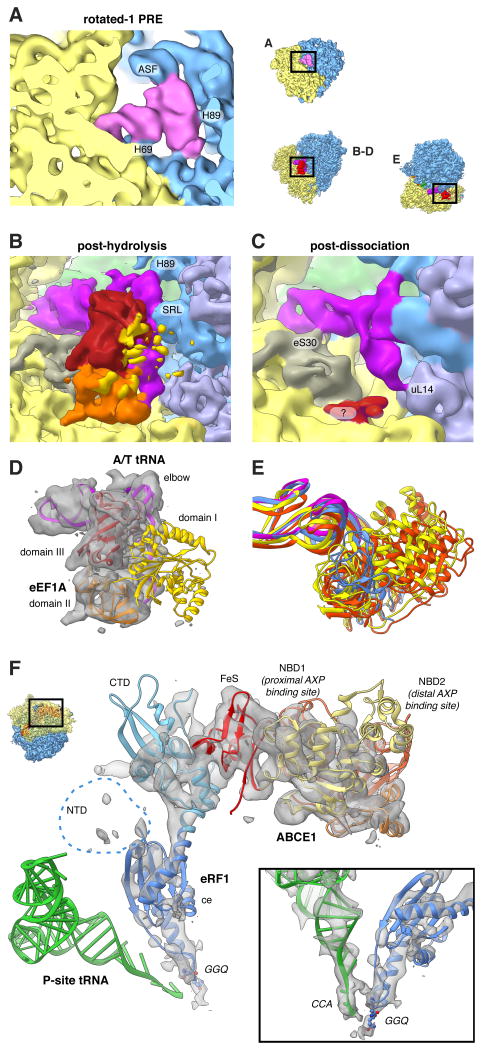
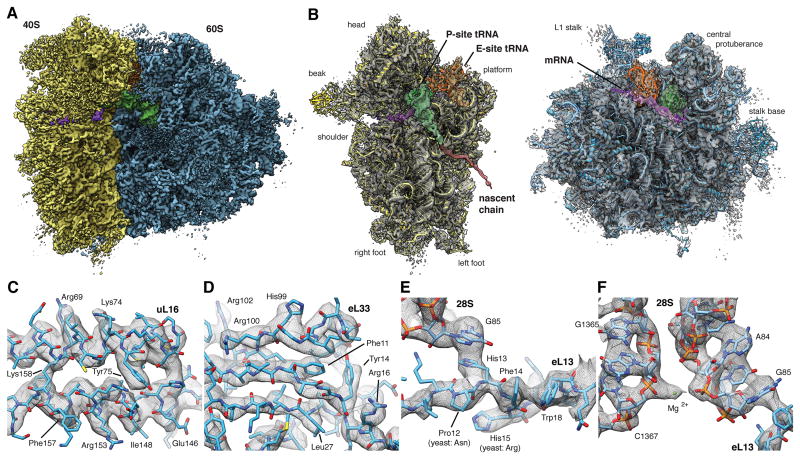

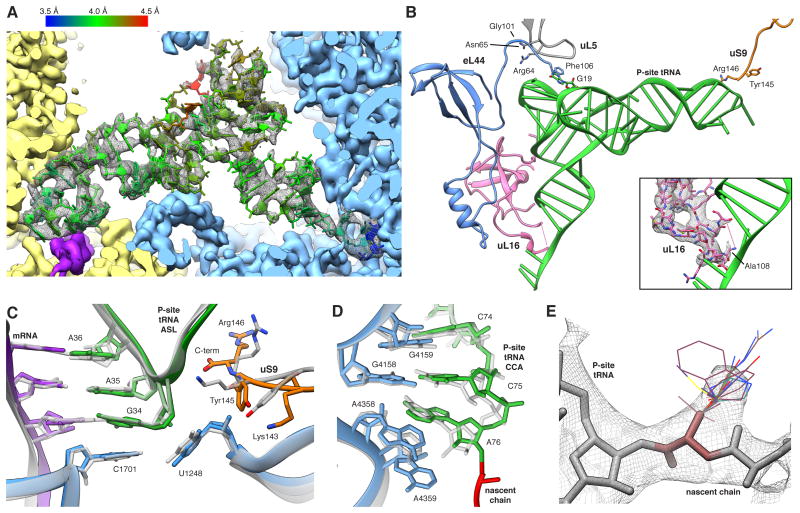
References
-
- Agmon I, Bashan A, Zarivach R, Yonath A. Symmetry at the active site of the ribosome: structural and functional implications. Biol Chem. 2005;386:833–844. - PubMed
-
- Anger AM, Armache JP, Berninghausen O, Habeck M, Subklewe M, Wilson DN, Beckmann R. Structures of the human and Drosophila 80S ribosome. Nature. 2013;497:80–85. - PubMed
-
- Ben-Shem A, Garreau de Loubresse N, Melnikov S, Jenner L, Yusupova G, Yusupov M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science. 2011;334:1524–1529. - PubMed
-
- Brandt F, Carlson LA, Hartl FU, Baumeister W, Griinewald K. The Three-Dimensional Organization of Polyribosomes in Intact Human Cells. Mol Cell. 2010;39:560–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
