

Co-transcriptional DNA and RNA Cleavage during Type III CRISPR-Cas Immunity
- PMID: 25959775
- PMCID: PMC4594840
- DOI: 10.1016/j.cell.2015.04.027
Co-transcriptional DNA and RNA Cleavage during Type III CRISPR-Cas Immunity
Abstract
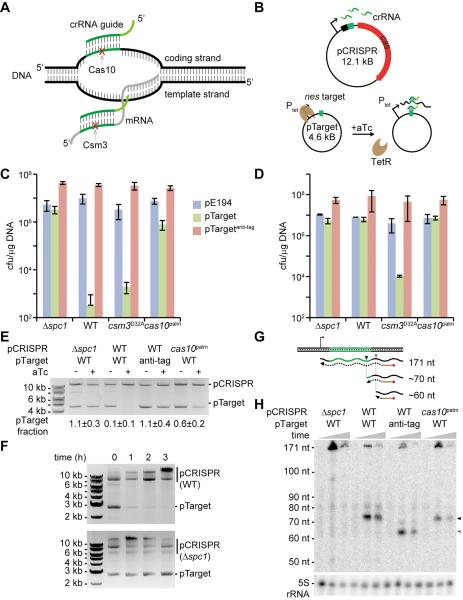
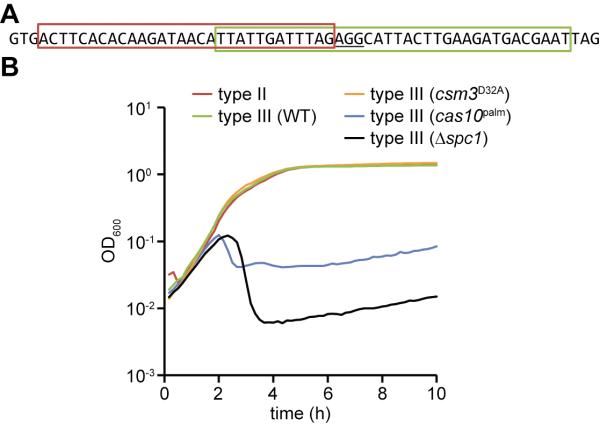
Immune systems must recognize and destroy different pathogens that threaten the host. CRISPR-Cas immune systems protect prokaryotes from viral and plasmid infection utilizing small CRISPR RNAs that are complementary to the invader's genome and specify the targets of RNA-guided Cas nucleases. Type III CRISPR-Cas immunity requires target transcription, and whereas genetic studies demonstrated DNA targeting, in vitro data have shown crRNA-guided RNA cleavage. The molecular mechanism behind these disparate activities is not known. Here, we show that transcription across the targets of the Staphylococcus epidermidis type III-A CRISPR-Cas system results in the cleavage of the target DNA and its transcripts, mediated by independent active sites within the Cas10-Csm ribonucleoprotein effector complex. Immunity against plasmids and DNA viruses requires DNA, but not RNA, cleavage activity. Our studies reveal a highly versatile mechanism of CRISPR immunity that can defend microorganisms against diverse DNA and RNA invaders.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
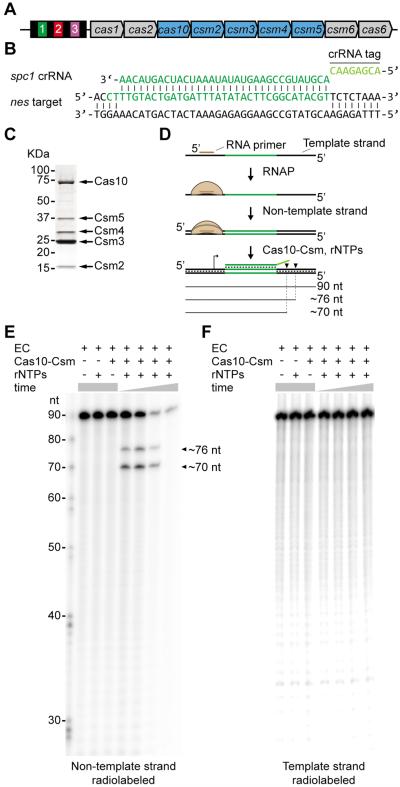
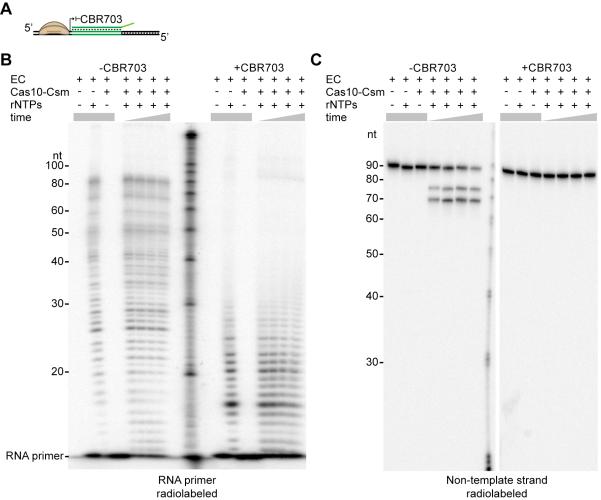

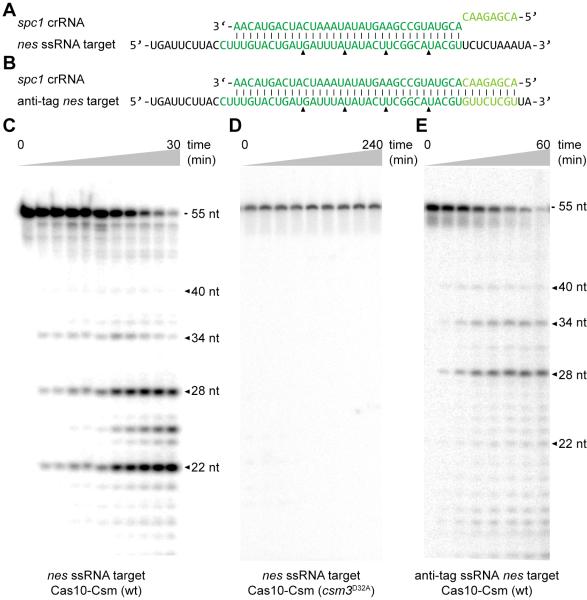
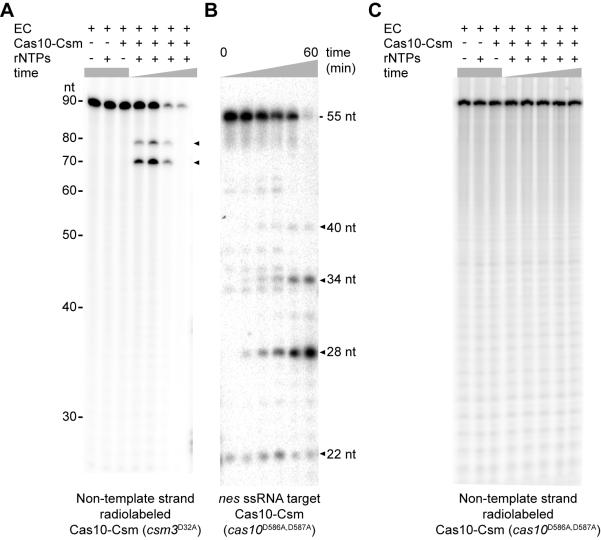
Comment in
-
A CRISPR View of Cleavage.Cell. 2015 May 21;161(5):964-966. doi: 10.1016/j.cell.2015.05.003. Cell. 2015. PMID: 26000476
References
-
- Artsimovitch I, Chu C, Lynch AS, Landick R. A new class of bacterial RNA polymerase inhibitor affects nucleotide addition. Science. 2003;302:650–654. - PubMed
-
- Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, Romero DA, Horvath P. CRISPR provides acquired resistance against viruses in prokaryotes. Science. 2007;315:1709–1712. - PubMed
-
- Bolotin A, Quinquis B, Sorokin A, Ehrlich SD. Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology. 2005;151:2551–2561. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
