

Direct Activation of STING in the Tumor Microenvironment Leads to Potent and Systemic Tumor Regression and Immunity
- PMID: 25959818
- PMCID: PMC4440852
- DOI: 10.1016/j.celrep.2015.04.031
Direct Activation of STING in the Tumor Microenvironment Leads to Potent and Systemic Tumor Regression and Immunity
Abstract
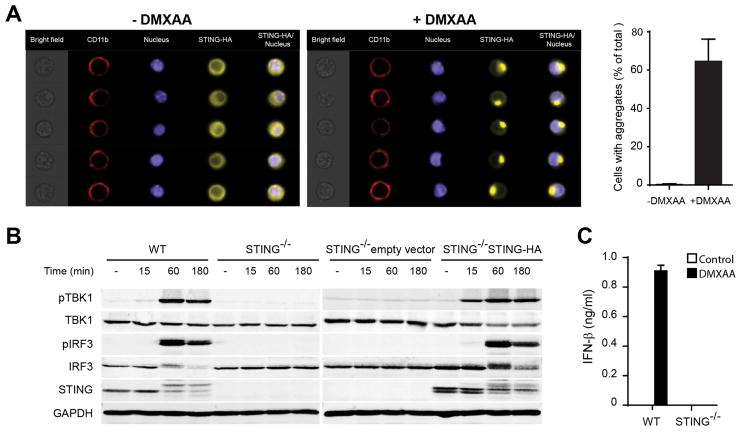
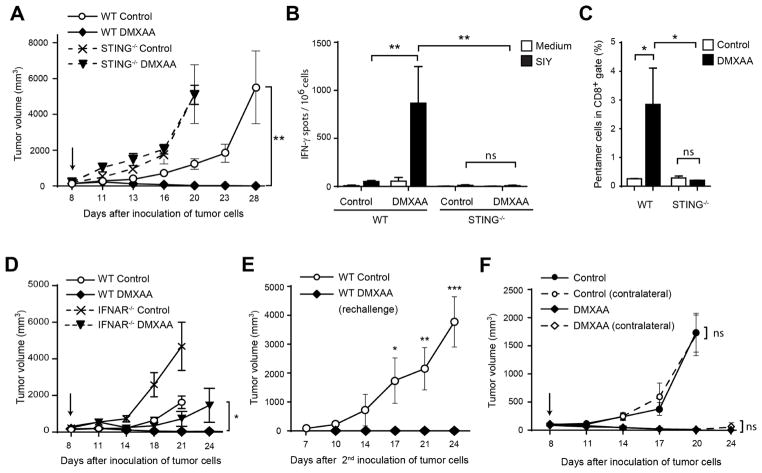
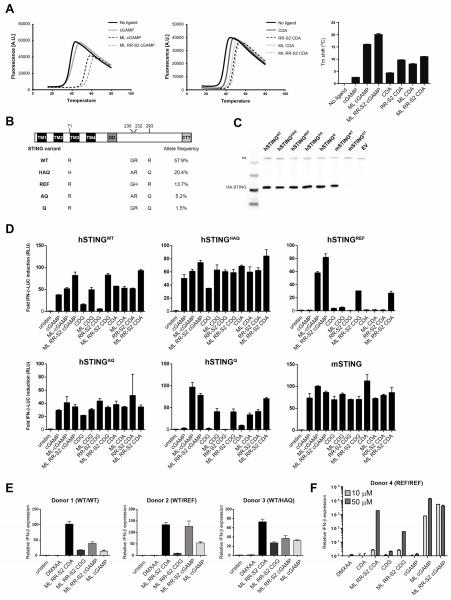
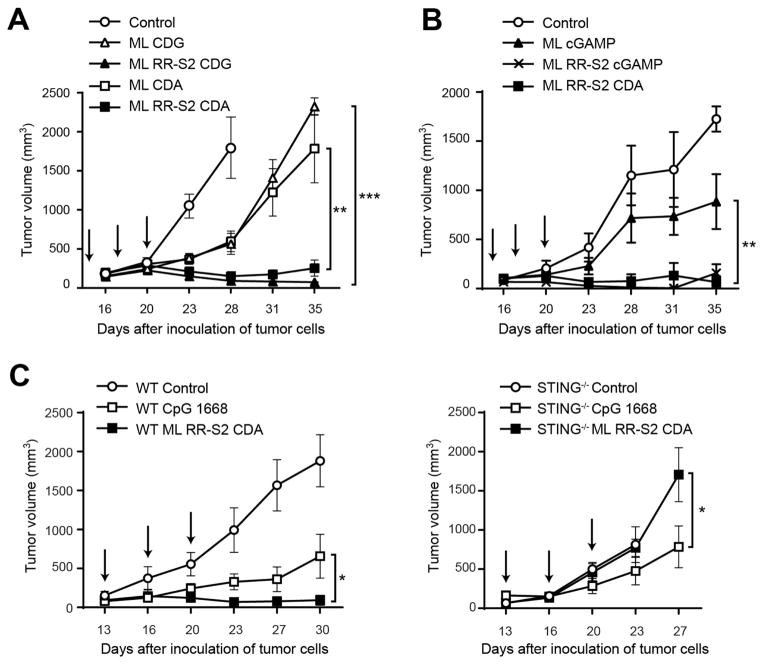
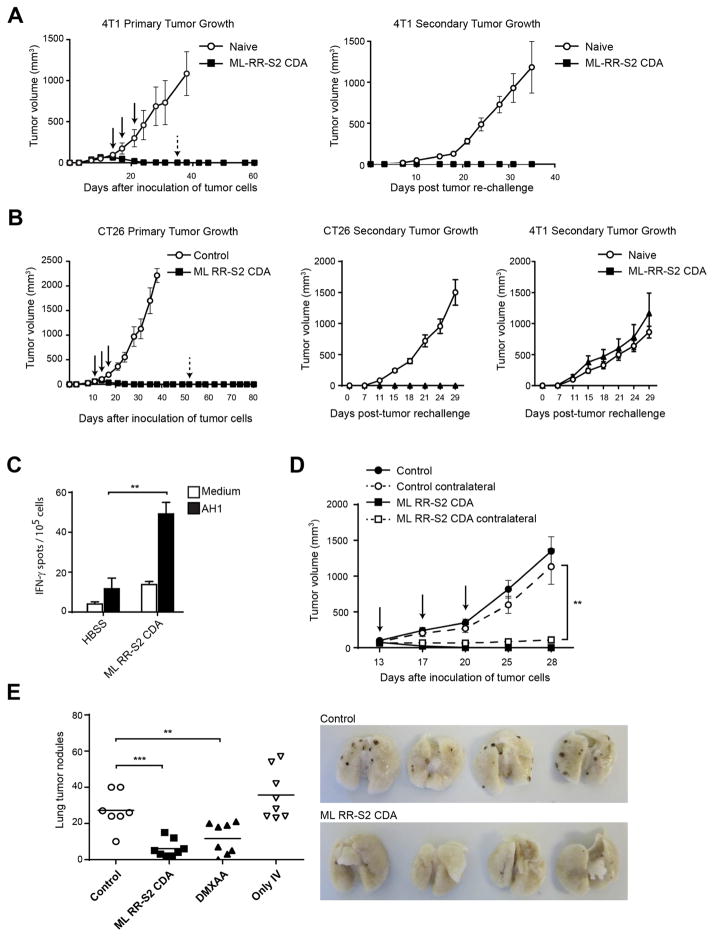
Spontaneous tumor-initiated T cell priming is dependent on IFN-β production by tumor-resident dendritic cells. On the basis of recent observations indicating that IFN-β expression was dependent upon activation of the host STING pathway, we hypothesized that direct engagement of STING through intratumoral (IT) administration of specific agonists would result in effective anti-tumor therapy. After proof-of-principle studies using the mouse STING agonist DMXAA showed a potent therapeutic effect, we generated synthetic cyclic dinucleotide (CDN) derivatives that activated all human STING alleles as well as murine STING. IT injection of STING agonists induced profound regression of established tumors in mice and generated substantial systemic immune responses capable of rejecting distant metastases and providing long-lived immunologic memory. Synthetic CDNs have high translational potential as a cancer therapeutic.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
L.H.G., S.M.M., D.B.K., K.E.S., G.E.K., E.L, T.B., J.J.L., K.M. and T.W.D are all paid employees of Aduro BioTech, hold stock in the company, and may be inventors on patent applications that apply to the CDN molecules described in the manuscript.
Figures
References
-
- Baguley BC, Ching LM. Immunomodulatory actions of xanthenone anticancer agents. BioDrugs: clinical immunotherapeutics, biopharmaceuticals and gene therapy. 1997;8:119–127. - PubMed
-
- Blank C, Brown I, Peterson AC, Spiotto M, Iwai Y, Honjo T, Gajewski TF. PD-L1/B7H-1 inhibits the effector phase of tumor rejection by T cell receptor (TCR) transgenic CD8+ T cells. Cancer Res. 2004;64:1140–1145. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
