

Structural basis for dynamic mechanism of nitrate/nitrite antiport by NarK
- PMID: 25959928
- PMCID: PMC4432589
- DOI: 10.1038/ncomms8097
Structural basis for dynamic mechanism of nitrate/nitrite antiport by NarK
Abstract
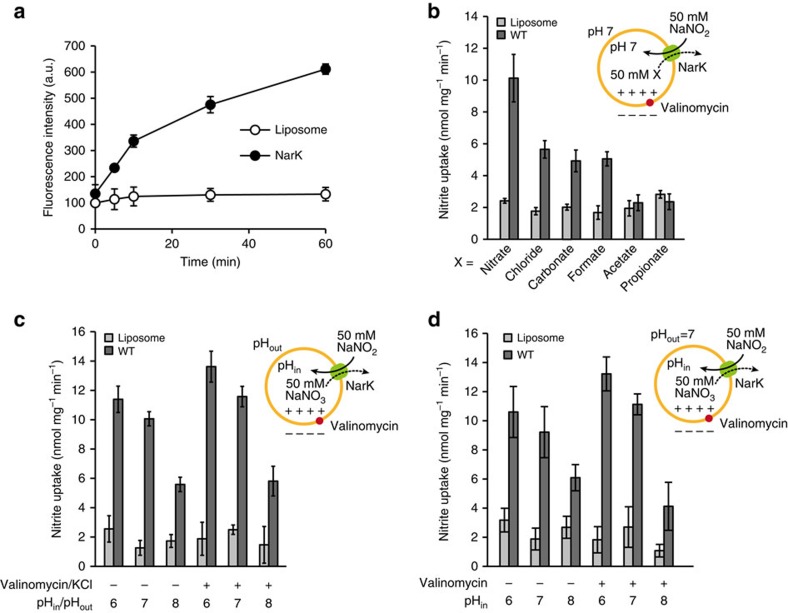
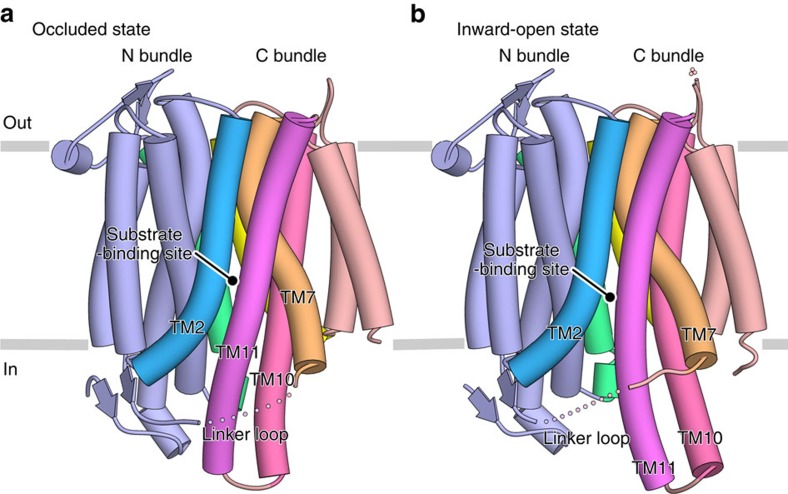
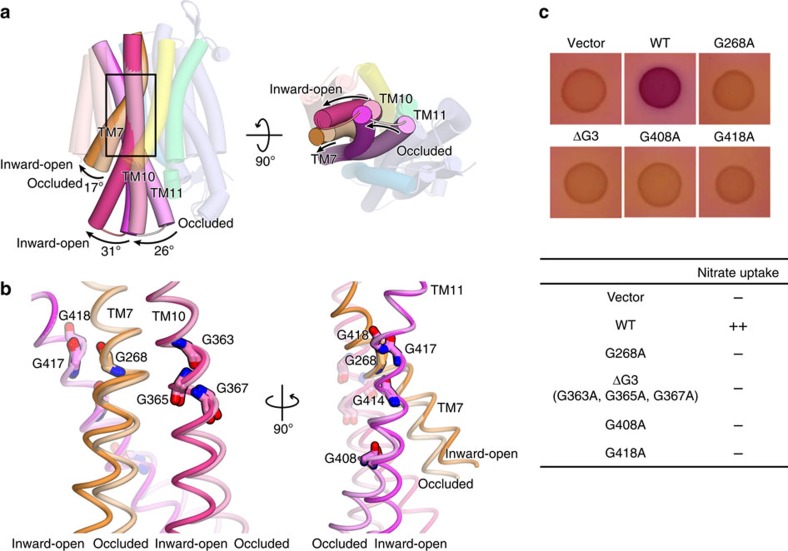
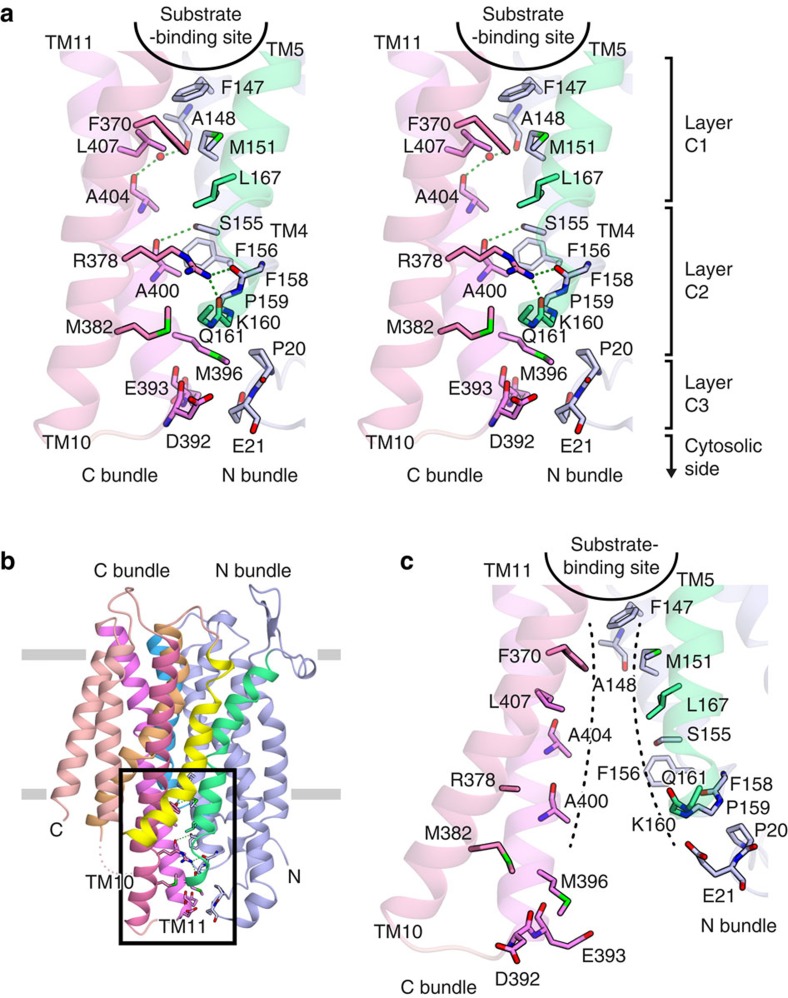
NarK belongs to the nitrate/nitrite porter (NNP) family in the major facilitator superfamily (MFS) and plays a central role in nitrate uptake across the membrane in diverse organisms, including archaea, bacteria, fungi and plants. Although previous studies provided insight into the overall structure and the substrate recognition of NarK, its molecular mechanism, including the driving force for nitrate transport, remained elusive. Here we demonstrate that NarK is a nitrate/nitrite antiporter, using an in vitro reconstituted system. Furthermore, we present the high-resolution crystal structures of NarK from Escherichia coli in the nitrate-bound occluded, nitrate-bound inward-open and apo inward-open states. The integrated structural, functional and computational analyses reveal the nitrate/nitrite antiport mechanism of NarK, in which substrate recognition is coupled to the transport cycle by the concomitant movement of the transmembrane helices and the key tyrosine and arginine residues in the substrate-binding site.
Figures
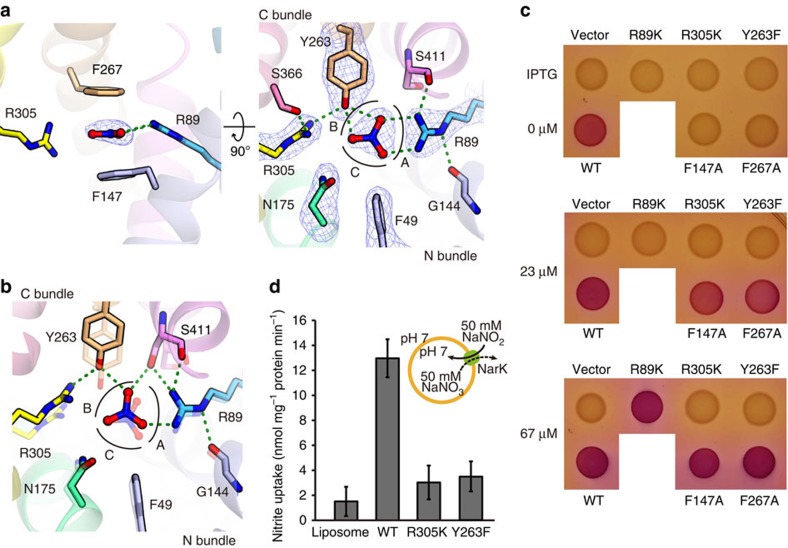
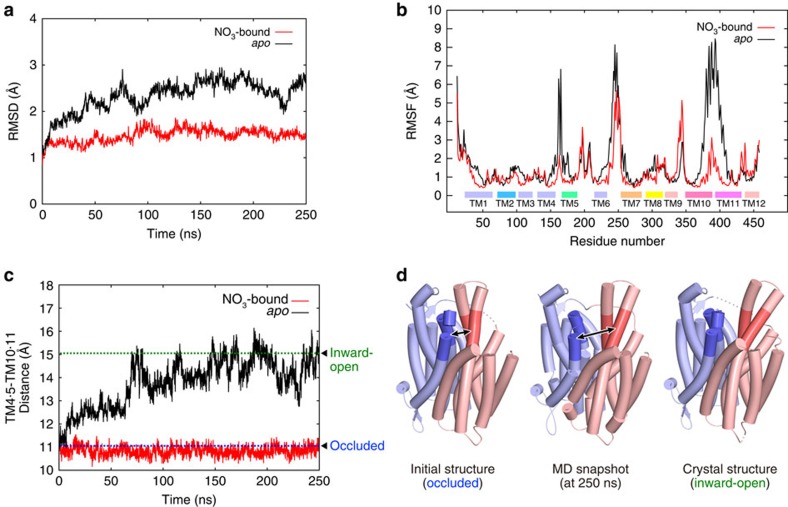
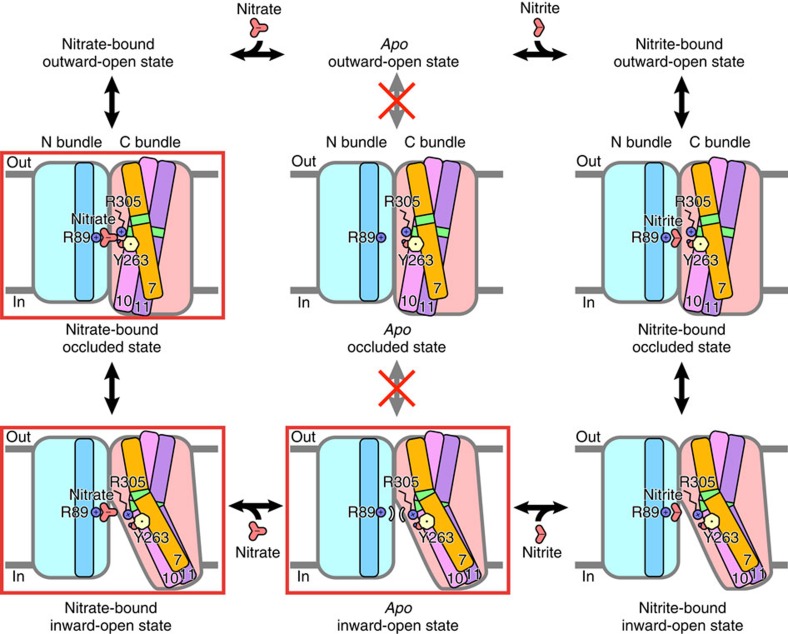
References
-
- Martínez-Espinosa R. M., Cole J. A., Richardson D. J. & Watmough N. J. Enzymology and ecology of the nitrogen cycle. Biochem. Soc. Trans. 39, 175–178 (2011). - PubMed
-
- Dixon R. & Kahn D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2, 621–631 (2004). - PubMed
-
- Nacry P., Bouguyon E. & Gojon A. Nitrogen acquisition by roots: physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil. 370, 1–29 (2013).
-
- Omata T. Structure, function and regulation of the nitrate transport system of the cyanobacterium Synechococcus sp. PCC7942. Plant Cell Physiol. 36, 207–213 (1995). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
