

Cryptochrome 1 regulates the circadian clock through dynamic interactions with the BMAL1 C terminus
- PMID: 25961797
- PMCID: PMC4456216
- DOI: 10.1038/nsmb.3018
Cryptochrome 1 regulates the circadian clock through dynamic interactions with the BMAL1 C terminus
Abstract
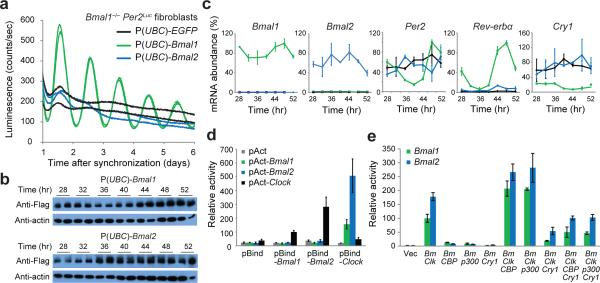
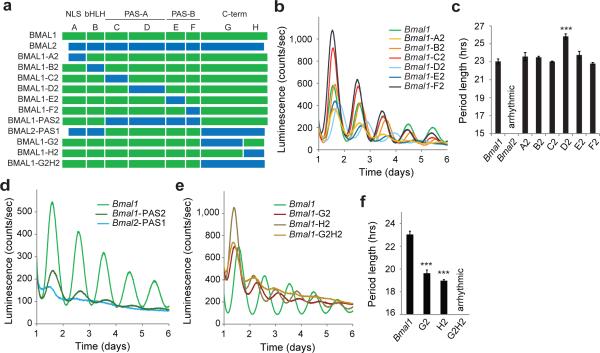
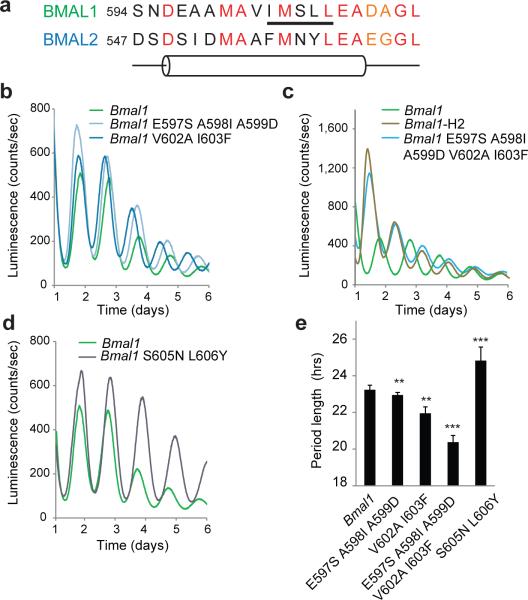
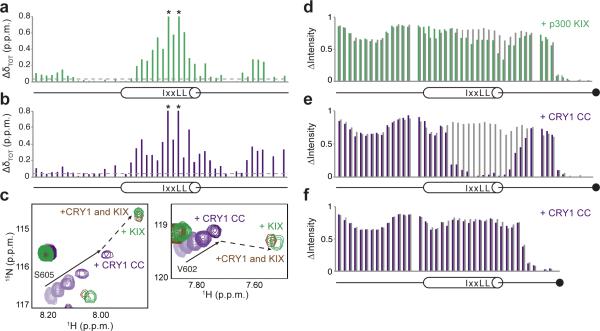
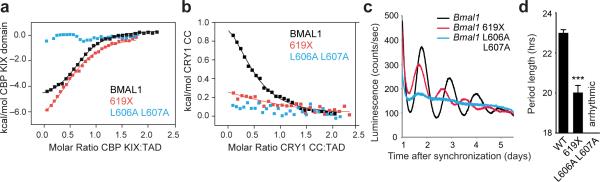
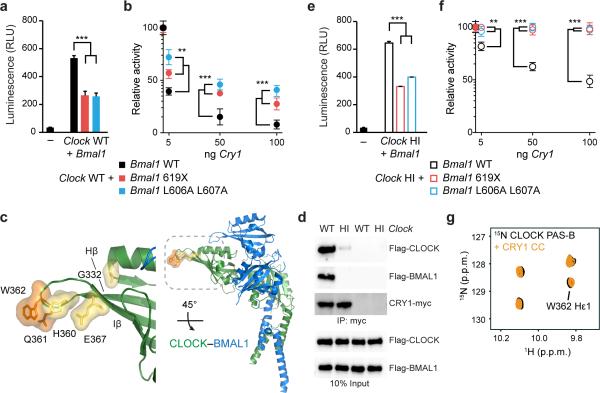
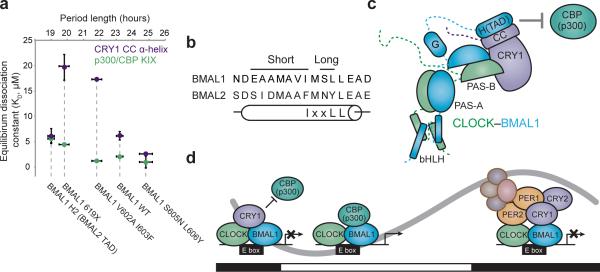
The molecular circadian clock in mammals is generated from transcriptional activation by the bHLH-PAS transcription factor CLOCK-BMAL1 and subsequent repression by PERIOD and CRYPTOCHROME (CRY). The mechanism by which CRYs repress CLOCK-BMAL1 to close the negative feedback loop and generate 24-h timing is not known. Here we show that, in mouse fibroblasts, CRY1 competes for binding with coactivators to the intrinsically unstructured C-terminal transactivation domain (TAD) of BMAL1 to establish a functional switch between activation and repression of CLOCK-BMAL1. TAD mutations that alter affinities for co-regulators affect the balance of repression and activation to consequently change the intrinsic circadian period or eliminate cycling altogether. Our results suggest that CRY1 fulfills its role as an essential circadian repressor by sequestering the TAD from coactivators, and they highlight regulation of the BMAL1 TAD as a critical mechanism for establishing circadian timing.
Figures
Comment in
-
Grab the wiggly tail: new insights into the dynamics of circadian clocks.Nat Struct Mol Biol. 2015 Jun;22(6):435-6. doi: 10.1038/nsmb.3039. Nat Struct Mol Biol. 2015. PMID: 26036566 No abstract available.
References
-
- Bass J. Circadian topology of metabolism. Nature. 2012;491:348–356. - PubMed
-
- Brown S, et al. PERIOD1-associated proteins modulate the negative limb of the mammalian circadian oscillator. Science. 2005;308:693–696. - PubMed
-
- Lee C, Etchegaray J, Cagampang F, Loudon A, Reppert S. Posttranslational mechanisms regulate the mammalian circadian clock. Cell. 2001;107:855–867. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
