

Metabolic regulation of hepatitis B immunopathology by myeloid-derived suppressor cells
- PMID: 25962123
- PMCID: PMC4458139
- DOI: 10.1038/nm.3856
Metabolic regulation of hepatitis B immunopathology by myeloid-derived suppressor cells
Abstract
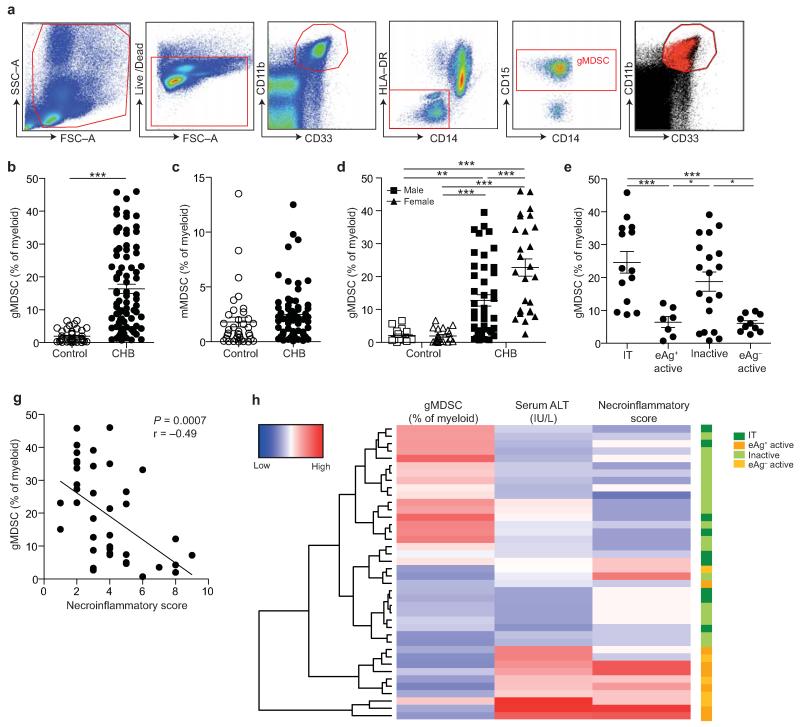
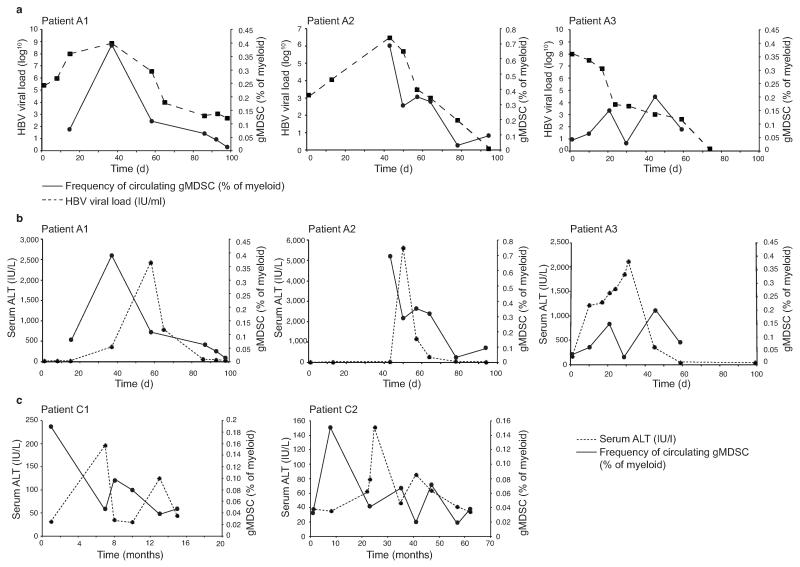
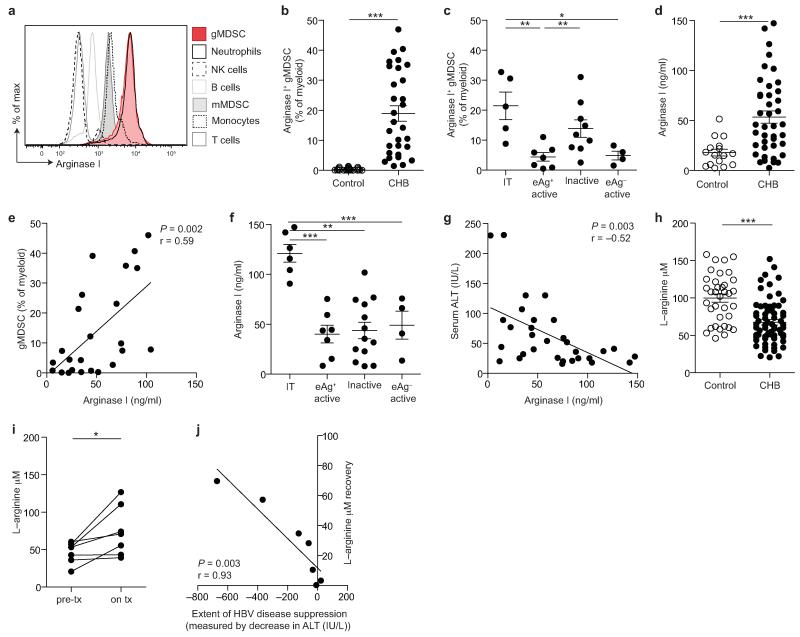
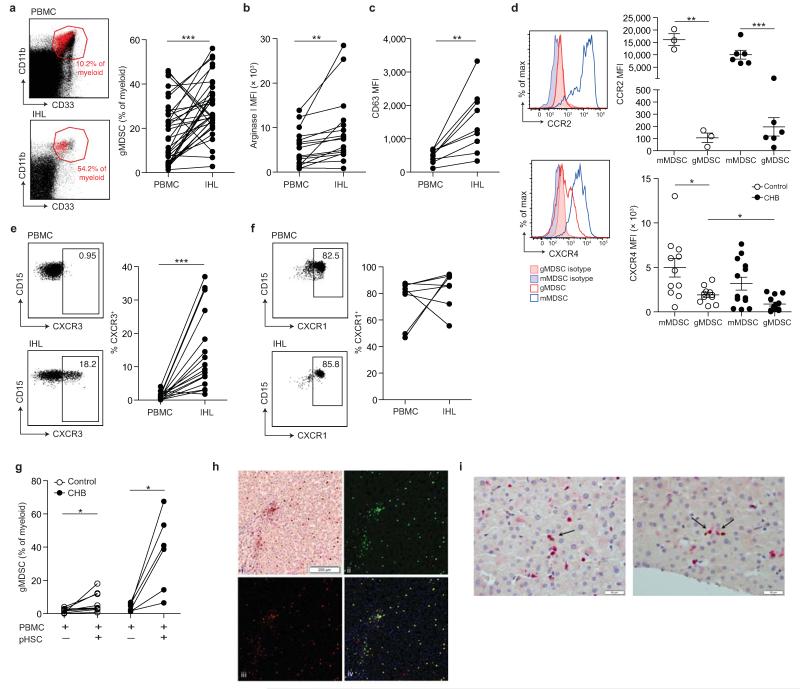
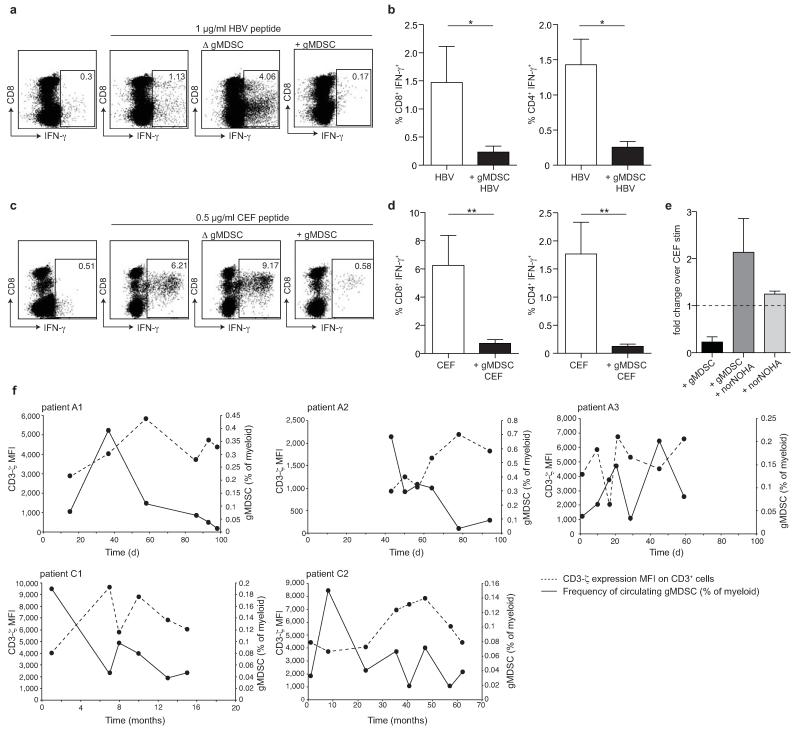
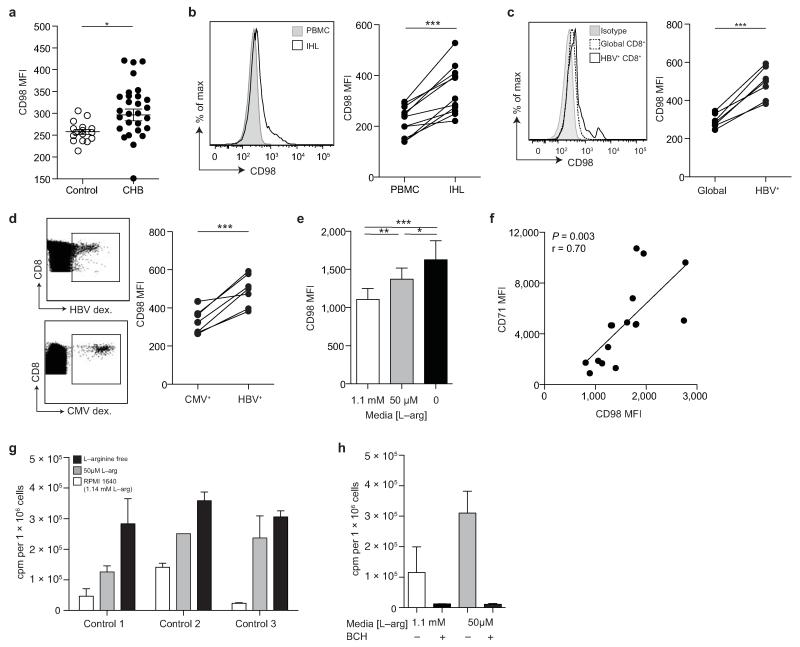
Infection with hepatitis B virus (HBV) results in disparate degrees of tissue injury: the virus can either replicate without pathological consequences or trigger immune-mediated necroinflammatory liver damage. We investigated the potential for myeloid-derived suppressor cells (MDSCs) to suppress T cell-mediated immunopathology in this setting. Granulocytic MDSCs (gMDSCs) expanded transiently in acute resolving HBV, decreasing in frequency prior to peak hepatic injury. In persistent infection, arginase-expressing gMDSCs (and circulating arginase) increased most in disease phases characterized by HBV replication without immunopathology, whilst L-arginine decreased. gMDSCs expressed liver-homing chemokine receptors and accumulated in the liver, their expansion supported by hepatic stellate cells. We provide in vitro and ex vivo evidence that gMDSCs potently inhibited T cells in a partially arginase-dependent manner. L-arginine-deprived T cells upregulated system L amino acid transporters to increase uptake of essential nutrients and attempt metabolic reprogramming. These data demonstrate the capacity of expanded arginase-expressing gMDSCs to regulate liver immunopathology in HBV infection.
Figures
Comment in
-
Hepatitis: Myeloid-derived suppressor cells in HBV infection.Nat Rev Gastroenterol Hepatol. 2015 Jul;12(7):370. doi: 10.1038/nrgastro.2015.89. Epub 2015 May 26. Nat Rev Gastroenterol Hepatol. 2015. PMID: 26008131 No abstract available.
-
Regulation of immunopathology in hepatitis B virus infection.Nat Med. 2015 Jun;21(6):548-9. doi: 10.1038/nm.3873. Nat Med. 2015. PMID: 26046570 No abstract available.
-
The challenge of protecting without overprotecting: The two sides of myeloid-derived suppressor cells in hepatitis B viral infection.Hepatology. 2016 Mar;63(3):1043-6. doi: 10.1002/hep.28385. Epub 2016 Jan 14. Hepatology. 2016. PMID: 26661682 No abstract available.
References
-
- Guidotti LG, Chisari FV. Immunobiology and pathogenesis of viral hepatitis. Annu Rev Pathol. 2006;1:23–61. - PubMed
-
- Bertoletti A, Maini MK. Protection or damage: a dual role for the virus-specific cytotoxic T lymphocyte response in hepatitis B and C infection? Curr Opin Immunol. 2000;12:403–408. - PubMed
-
- Kakimi K, et al. Blocking chemokine responsive to gamma-2/interferon (IFN)-gamma inducible protein and monokine induced by IFN-gamma activity in vivo reduces the pathogenetic but not the antiviral potential of hepatitis B virus-specific cytotoxic T lymphocytes. J Exp Med. 2001;194:1755–1766. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
