

Pathogenicity of Highly Pathogenic Avian Influenza Virus H5N1 in Naturally Infected Poultry in Egypt
- PMID: 25962145
- PMCID: PMC4427178
- DOI: 10.1371/journal.pone.0120061
Pathogenicity of Highly Pathogenic Avian Influenza Virus H5N1 in Naturally Infected Poultry in Egypt
Abstract
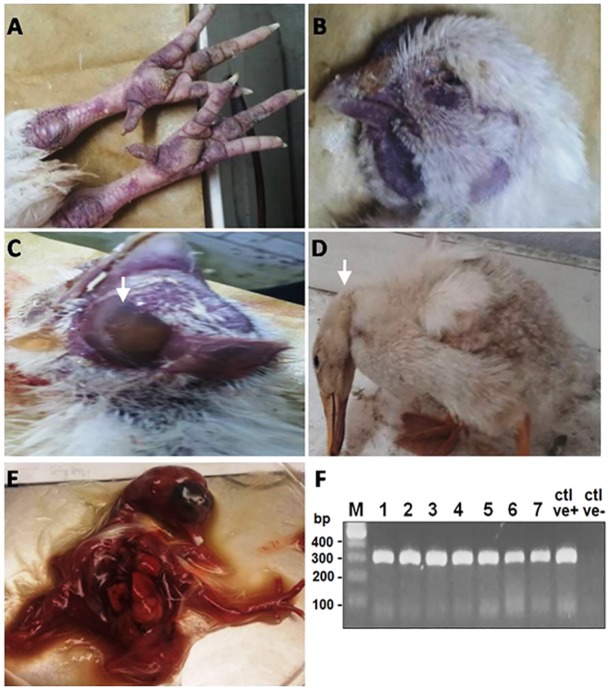
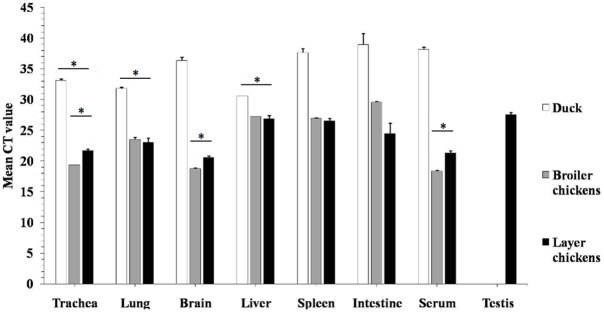
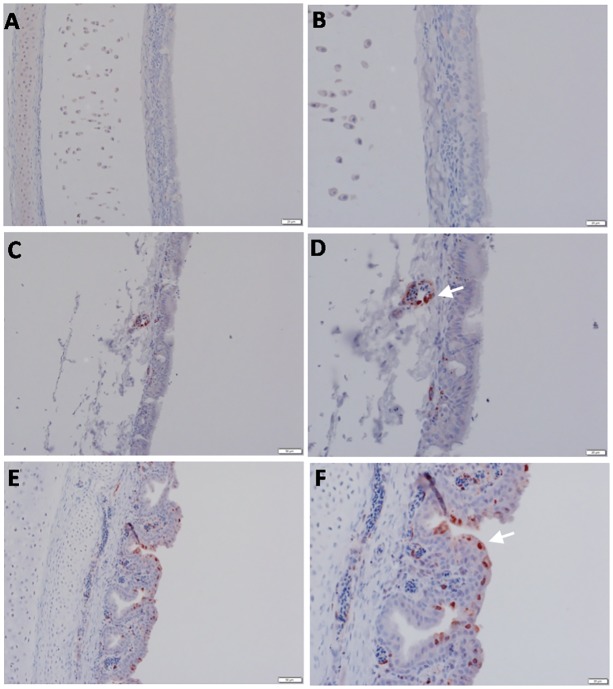
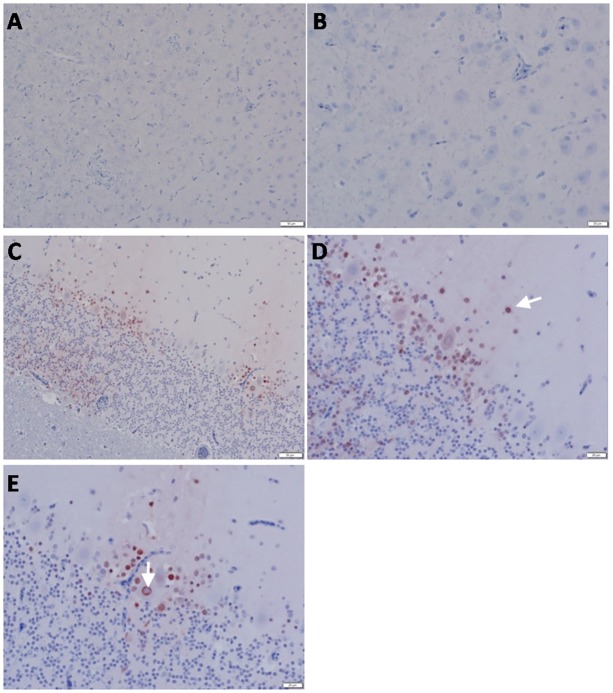
Highly pathogenic avian influenza virus (HPAIV) H5N1 has been endemic in Egypt since 2006, and there is increasing concern for its potential to become highly transmissible among humans. Infection by HPAIV H5N1 has been described in experimentally challenged birds. However, the pathogenicity of the H5N1 isolated in Egypt has never been reported in naturally infected chickens and ducks. Here we report a 2013 outbreak of HPAIV H5N1 in commercial poultry farms and backyards in Sharkia Province, Egypt. The main symptoms were ecchymosis on the shanks and feet, cyanosis of the comb and wattles, subcutaneous edema of the head and neck for chickens, and nervous signs (torticollis) for ducks. Within 48-72 hrs of the onset of illness, the average mortality rates were 22.8-30% and 28.5-40% in vaccinated chickens and non-vaccinated ducks, respectively. Tissue samples of chickens and ducks were collected for analyses with cross-section immunohistochemistry and real-time RT-PCR for specific viral RNA transcripts. While viral RNA was detected in nearly all tissues and sera collected, viral nucleoprotein was detected almost ubiquitously in all tissues, including testis. Interestingly, viral antigen was also observed in endothelial cells of most organs in chickens, and clearly detected in the trachea and brain in particular. Viral nucleoprotein was also detected in mononuclear cells of various organs, especially pulmonary tissue. We performed phylogenetic analyses and compared the genomic sequences of the hemagglutinin (HA) and nonstructural proteins (NS) among the isolated viruses, the HPAIV circulated in Egypt in the past and currently, and some available vaccine strains. Further analysis of deduced amino acids of both HA and NS1 revealed that our isolates carried molecular determinants of HPAIV, including the multibasic amino acids (PQGERRRK/KR*GLF) in the cleavage site in HA and glutamate at position 92 (D92E) in NS1. This is the first report of the pathogenicity of the HPAIVH5N1 strain currently circulating in naturally infected poultry in Egypt, which may provide unique insights into the viral pathogenesis in HPAIV-infected chickens and ducks.
Conflict of interest statement
Figures
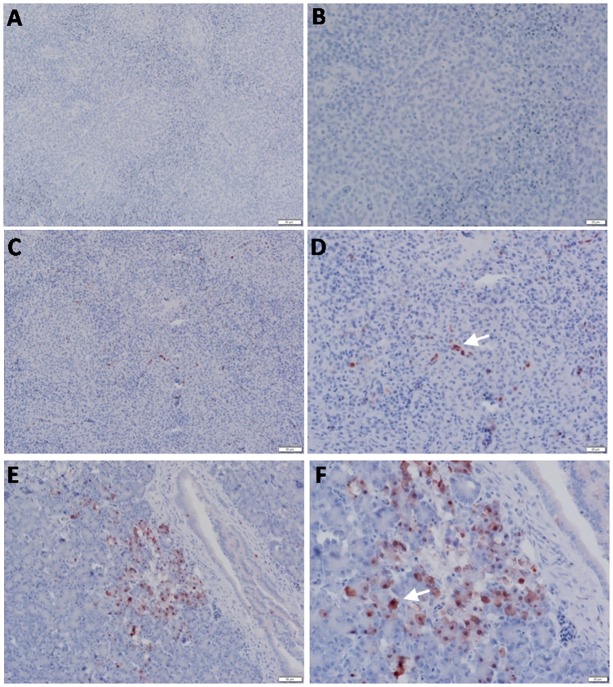
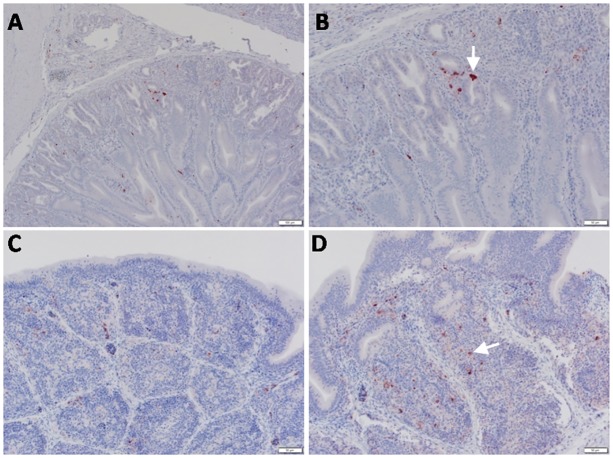
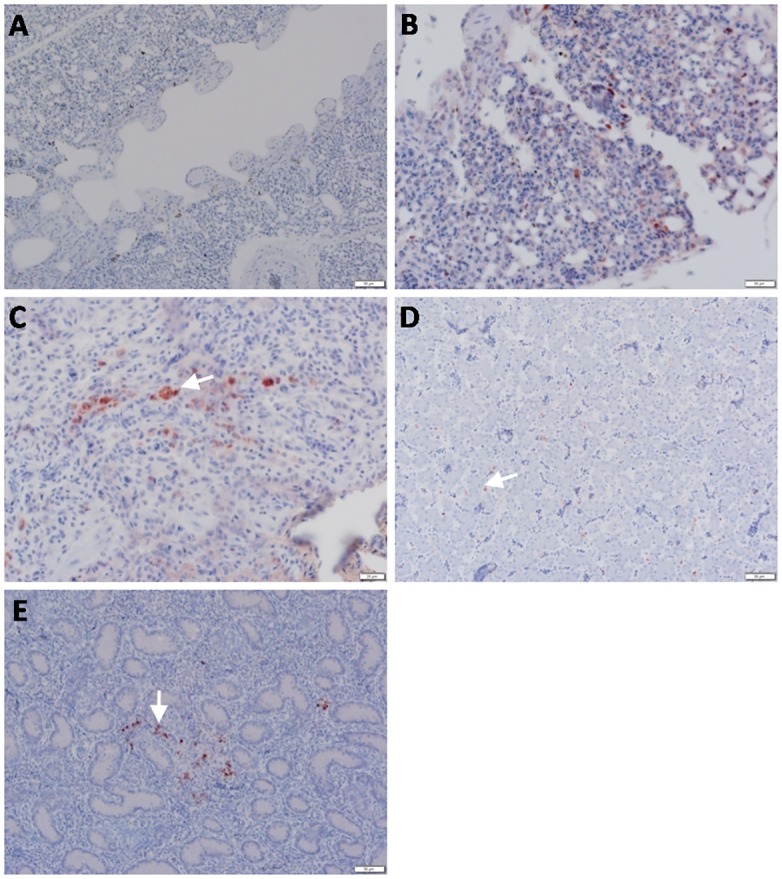

References
-
- Xu X, Subbarao K, Cox NJ, Guo Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology. 1999;261(1):15–9. - PubMed
-
- Sims L, Ellis T, Liu K, Dyrting K, Wong H, Peiris M, et al. Avian influenza in Hong Kong 1997–2002. Avian diseases. 2003;47(s3):832–8. - PubMed
-
- Sims L, Domenech J, Benigno C, Kahn S, Kamata A, Lubroth J, et al. Origin and evolution of highly pathogenic H5N1 avian influenza in Asia. Veterinary Record. 2005;157(6):159 - PubMed
-
- Chen H, Smith G, Zhang S, Qin K, Wang J, Li K, et al. Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature. 2005;436(7048):191–2. - PubMed
-
- Liu J, Xiao H, Lei F, Zhu Q, Qin K, Zhang X-w, et al. Highly pathogenic H5N1 influenza virus infection in migratory birds. Science. 2005;309(5738):1206-. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
