

Tomato plant cell death induced by inhibition of HSP90 is alleviated by Tomato yellow leaf curl virus infection
- PMID: 25962748
- PMCID: PMC6638530
- DOI: 10.1111/mpp.12275
Tomato plant cell death induced by inhibition of HSP90 is alleviated by Tomato yellow leaf curl virus infection
Abstract
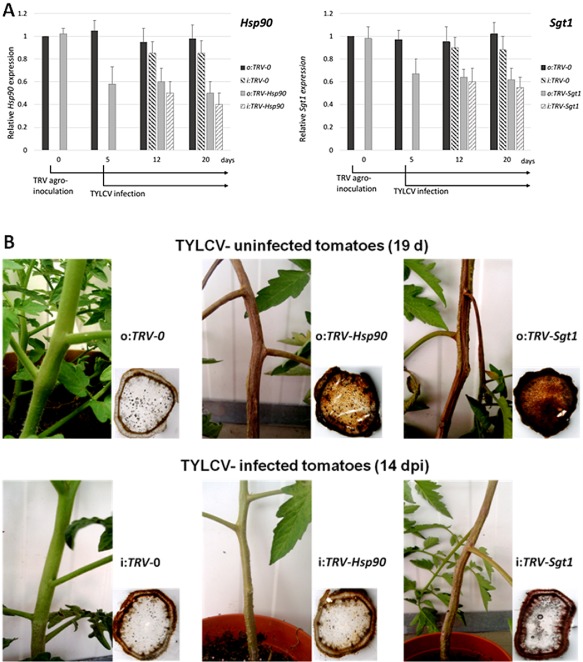
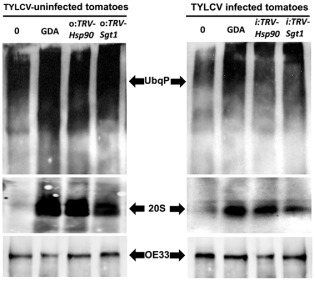
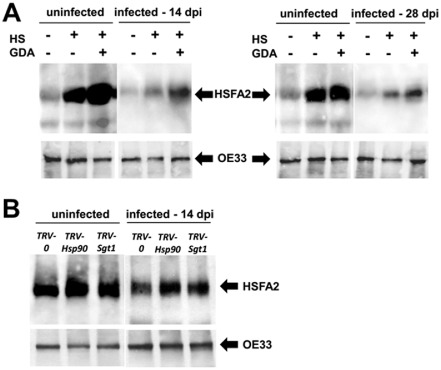
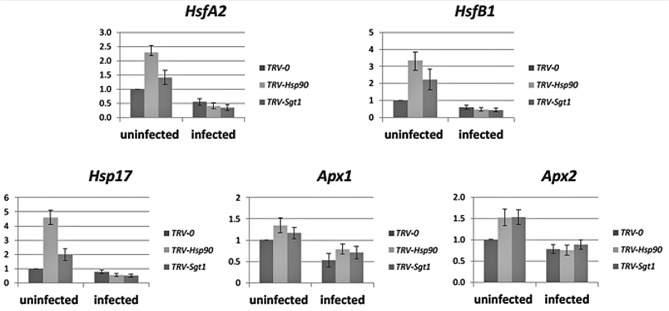
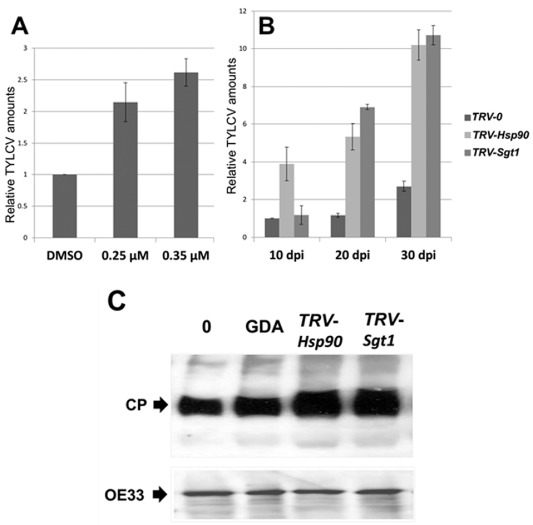
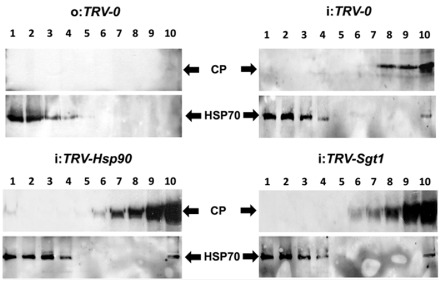
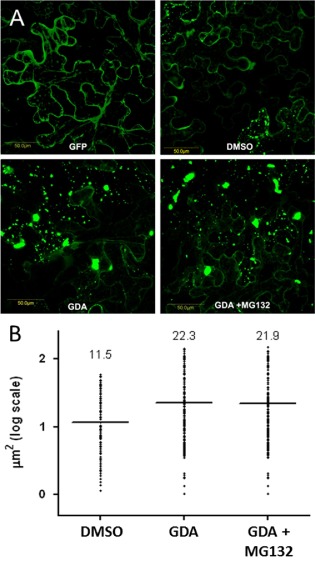
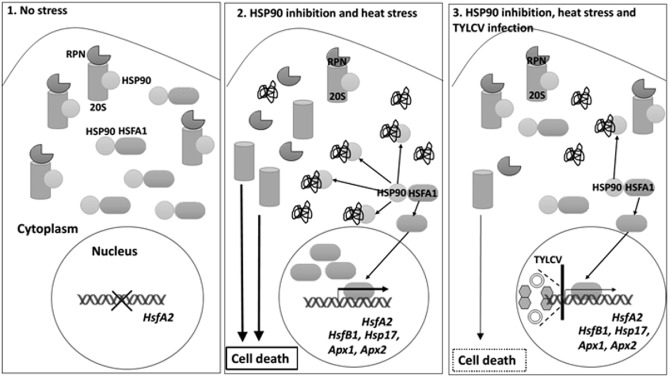
To ensure a successful long-term infection cycle, begomoviruses must restrain their destructive effect on host cells and prevent drastic plant responses, at least in the early stages of infection. The monopartite begomovirus Tomato yellow leaf curl virus (TYLCV) does not induce a hypersensitive response and cell death on whitefly-mediated infection of virus-susceptible tomato plants until diseased tomatoes become senescent. The way in which begomoviruses evade plant defences and interfere with cell death pathways is still poorly understood. We show that the chaperone HSP90 (heat shock protein 90) and its co-chaperone SGT1 (suppressor of the G2 allele of Skp1) are involved in the establishment of TYLCV infection. Inactivation of HSP90, as well as silencing of the Hsp90 and Sgt1 genes, leads to the accumulation of damaged ubiquitinated proteins and to a cell death phenotype. These effects are relieved under TYLCV infection. HSP90-dependent inactivation of 26S proteasome degradation and the transcriptional activation of the heat shock transcription factors HsfA2 and HsfB1 and of the downstream genes Hsp17 and Apx1/2 are suppressed in TYLCV-infected tomatoes. Following suppression of the plant stress response, TYLCV can replicate and accumulate in a permissive environment.
Keywords: HSP90; cell death; geminivirus; heat stress transcription factor.
© 2015 BSPP AND JOHN WILEY & SONS LTD.
Figures
References
-
- Alvarez, M.E. , Pennell, R.I. , Meijer, P.J. , Ishikawa, A. , Dixon, R.A. and Lamb, C. (1998) Reactive oxygen intermediates mediate a systemic signal network in the establishment of plant immunity. Cell, 92, 773–784. - PubMed
-
- Ascencio‐Ibanez, J.T. , Sozzani, R. , Lee, T.J. , Chu, T.M. , Wolfinger, R.D. , Cella, R. and Hanley‐Bowdoin, L. (2008) Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 148, 436–454. - PMC - PubMed
-
- Baniwal, S.K. , Bharti, K. , Chan, K.Y. , Fauth, M. , Ganguli, A. , Kotak, S. , Mishra, S.K. , Nover, L. , Port, M. , Scharf, K.D. , Tripp, J. , Weber, C. , Zielinski, D. and von Koskull‐Döring, P. (2004) Heat stress response in plants: a complex game with chaperones and more than twenty heat stress transcription factors. J. Biosci. 29, 471–487. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
