

Mitochondrial DNA variants can mediate methylation status of inflammation, angiogenesis and signaling genes
- PMID: 25964427
- PMCID: PMC4512622
- DOI: 10.1093/hmg/ddv173
Mitochondrial DNA variants can mediate methylation status of inflammation, angiogenesis and signaling genes
Abstract
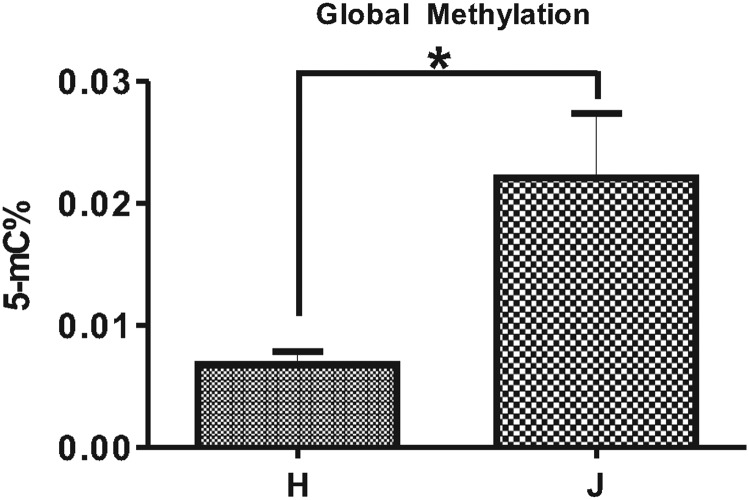
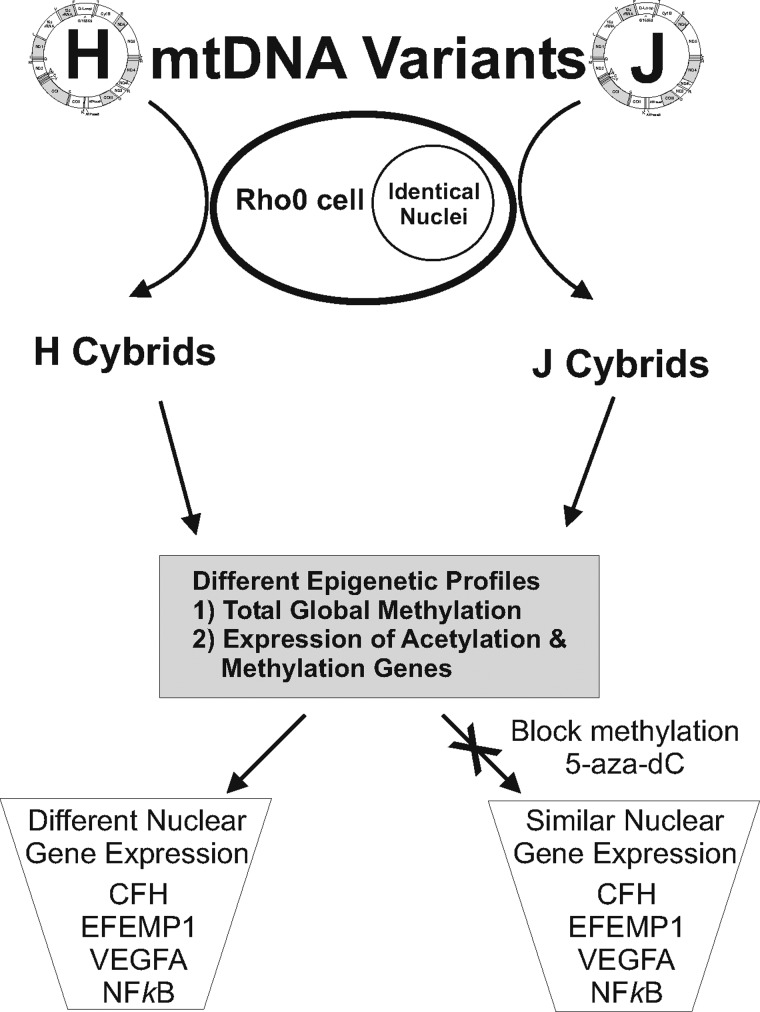
Mitochondrial (mt) DNA can be classified into haplogroups representing different geographic and/or racial origins of populations. The H haplogroup is protective against age-related macular degeneration (AMD), while the J haplogroup is high risk for AMD. In the present study, we performed comparison analyses of human retinal cell cybrids, which possess identical nuclei, but mtDNA from subjects with either the H or J haplogroups, and demonstrate differences in total global methylation, and expression patterns for two genes related to acetylation and five genes related to methylation. Analyses revealed that untreated-H and -J cybrids have different expression levels for nuclear genes (CFH, EFEMP1, VEGFA and NFkB2). However, expression levels for these genes become equivalent after treatment with a methylation inhibitor, 5-aza-2'-deoxycytidine. Moreover, sequencing of the entire mtDNA suggests that differences in epigenetic status found in cybrids are likely due to single nucleotide polymorphisms (SNPs) within the haplogroup profiles rather than rare variants or private SNPs. In conclusion, our findings indicate that mtDNA variants can mediate methylation profiles and transcription for inflammation, angiogenesis and various signaling pathways, which are important in several common diseases.
© The Author 2015. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures

References
-
- Wallace D.C., Lott M.T., Procaccio V. (2007) Mitochondrial genes in degenerative diseases, cancer and aging. In: Rimoin C., Pyeritz K. (eds), Emery and Rimoin's Principles and Practices of Medical Genetics. Churchill Livingstone Elsevier, Philadelphia, PA.
-
- Jones M.M., Manwaring N., Wang J.J., Rochtchina E., Mitchell P., Sue C.M. (2007) Mitochondrial DNA haplogroups and age-related maculopathy. Arch. Ophthalmol., 125, 1235–1240. - PubMed
-
- Udar N., Atilano S.R., Memarzadeh M., Boyer D., Chwa M., Lu S., Maguen B., Langberg J., Coskun P., Wallace D.C., et al. (2009) Mitochondrial DNA haplogroups associated with age-related macular degeneration. Invest. Ophthalmol. Vis. Sci., 50, 2966–2974. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
