

Plasma Membrane Proteolipid 3 Protein Modulates Amphotericin B Resistance through Sphingolipid Biosynthetic Pathway
- PMID: 25965669
- PMCID: PMC4428271
- DOI: 10.1038/srep09685
Plasma Membrane Proteolipid 3 Protein Modulates Amphotericin B Resistance through Sphingolipid Biosynthetic Pathway
Abstract
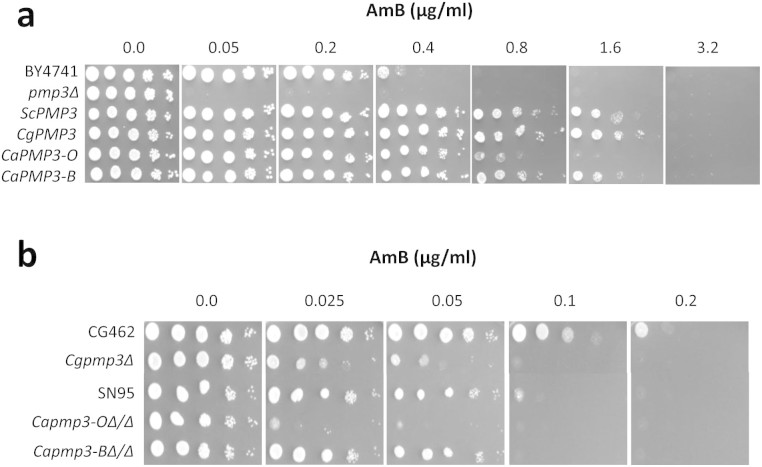
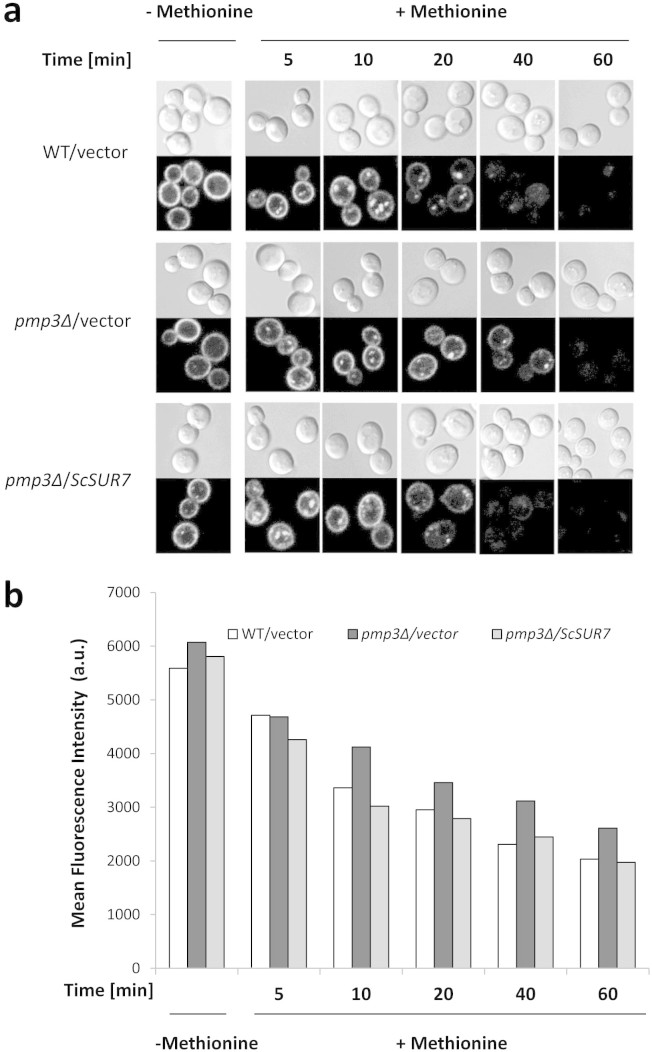
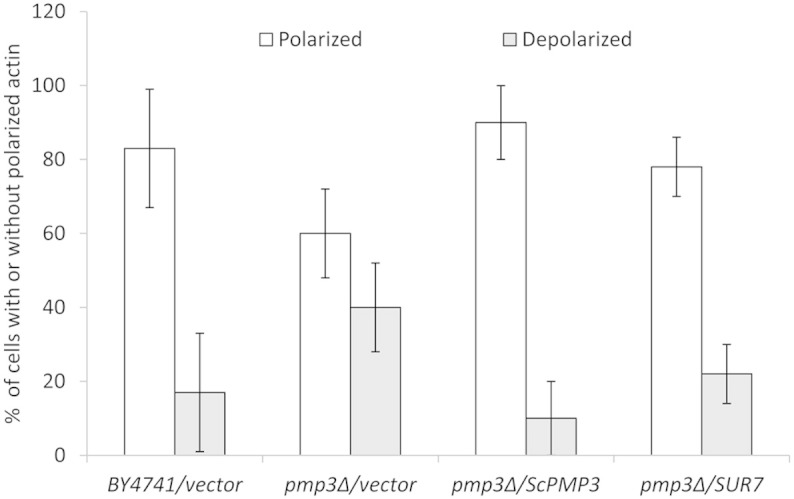
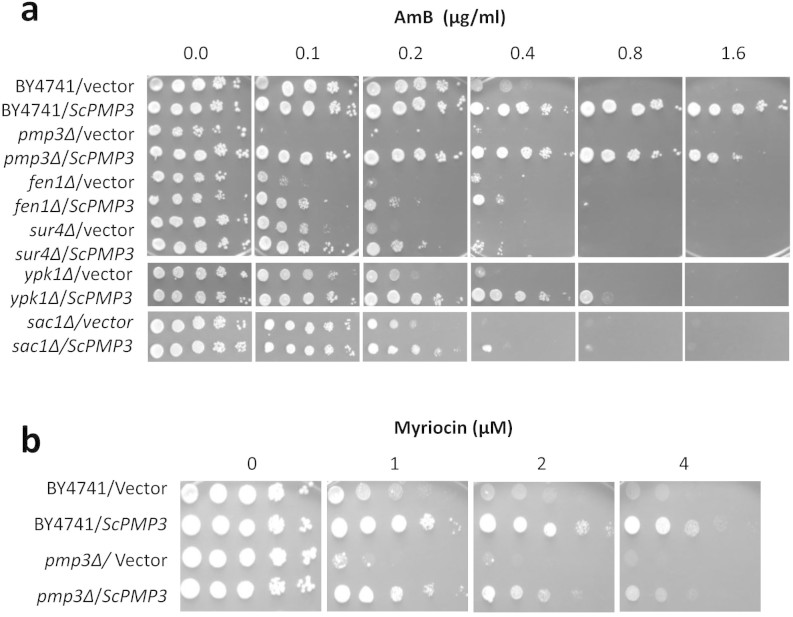
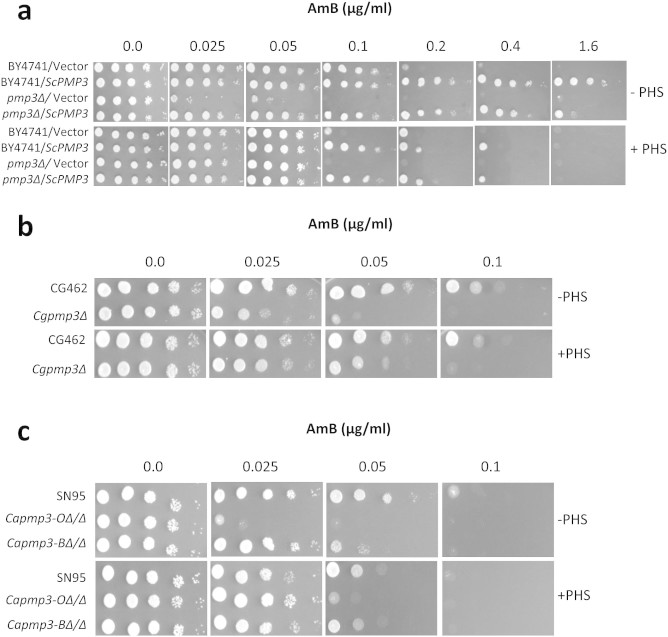
Invasive opportunistic fungal infections of humans are common among those suffering from impaired immunity, and are difficult to treat resulting in high mortality. Amphotericin B (AmB) is one of the few antifungals available to treat such infections. The AmB resistance mechanisms reported so far mainly involve decrease in ergosterol content or alterations in cell wall. In contrast, depletion of sphingolipids sensitizes cells to AmB. Recently, overexpression of PMP3 gene, encoding plasma membrane proteolipid 3 protein, was shown to increase and its deletion to decrease, AmB resistance. Here we have explored the mechanistic basis of PMP3 effect on AmB resistance. It was found that ergosterol content and cell wall integrity are not related to modulation of AmB resistance by PMP3. A few prominent phenotypes of PMP3 delete strain, namely, defective actin polarity, impaired salt tolerance, and reduced rate of endocytosis are also not related to its AmB-sensitivity. However, PMP3 overexpression mediated increase in AmB resistance requires a functional sphingolipid pathway. Moreover, AmB sensitivity of strains deleted in PMP3 can be suppressed by the addition of phytosphingosine, a sphingolipid pathway intermediate, confirming the importance of this pathway in modulation of AmB resistance by PMP3.
Figures

References
-
- Brown G. D. et al. Hidden killers: human fungal infections. Sci Transl Med 4, 165rv113 (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
