

Bacterial swarms recruit cargo bacteria to pave the way in toxic environments
- PMID: 25968641
- PMCID: PMC4436059
- DOI: 10.1128/mBio.00074-15
Bacterial swarms recruit cargo bacteria to pave the way in toxic environments
Abstract
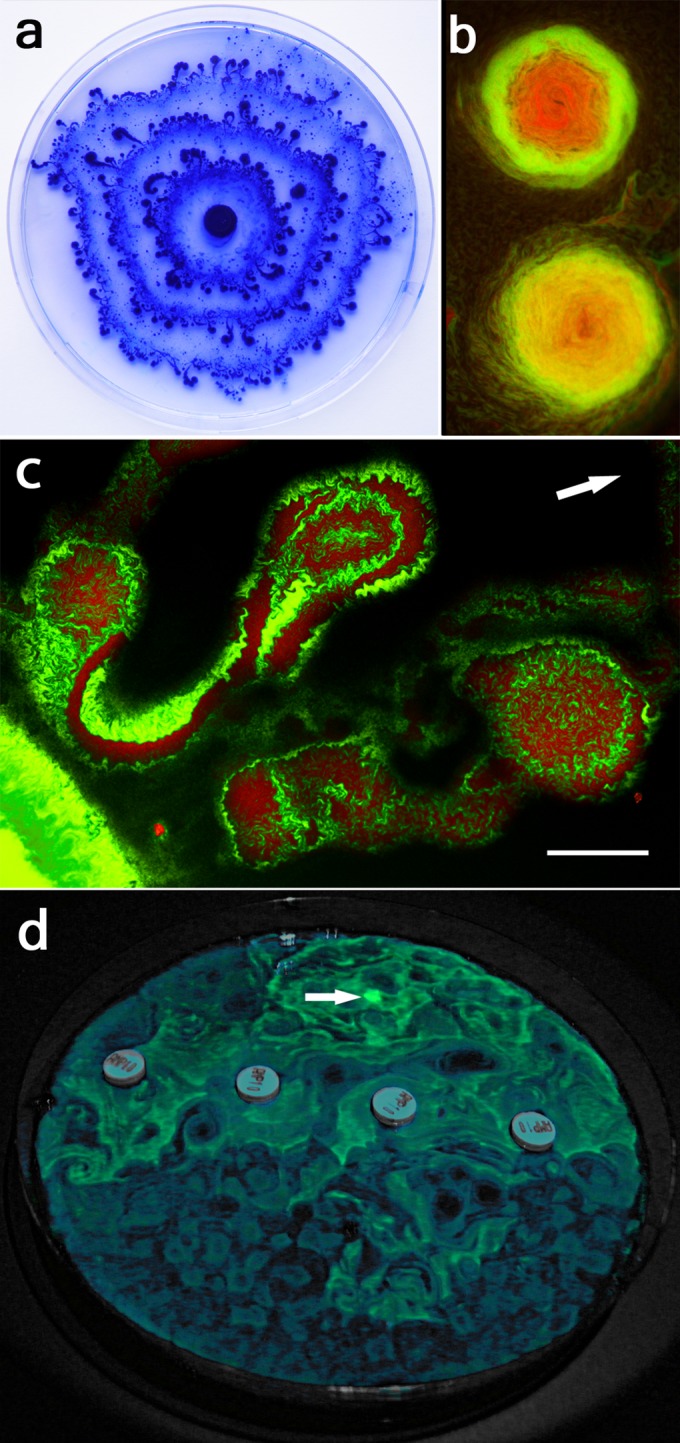
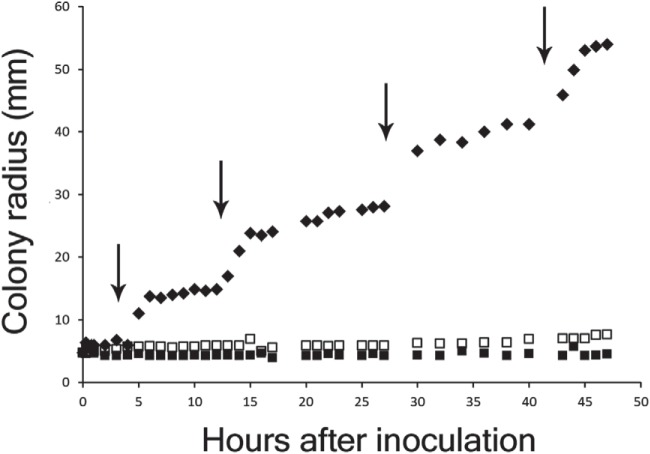
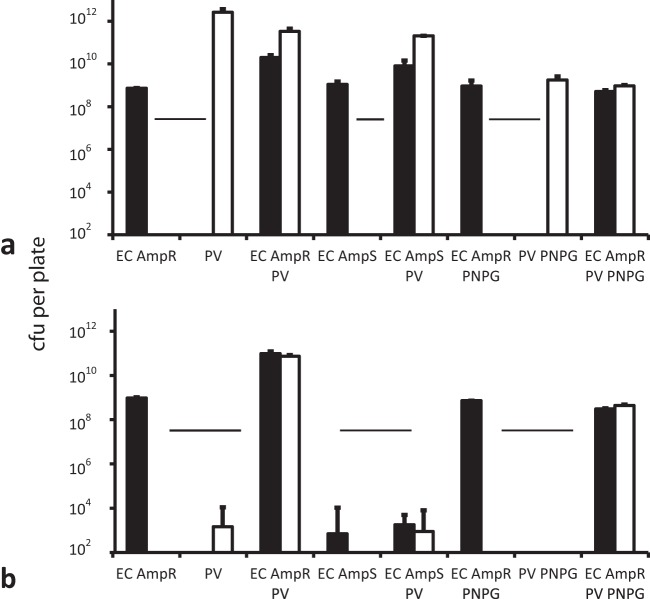
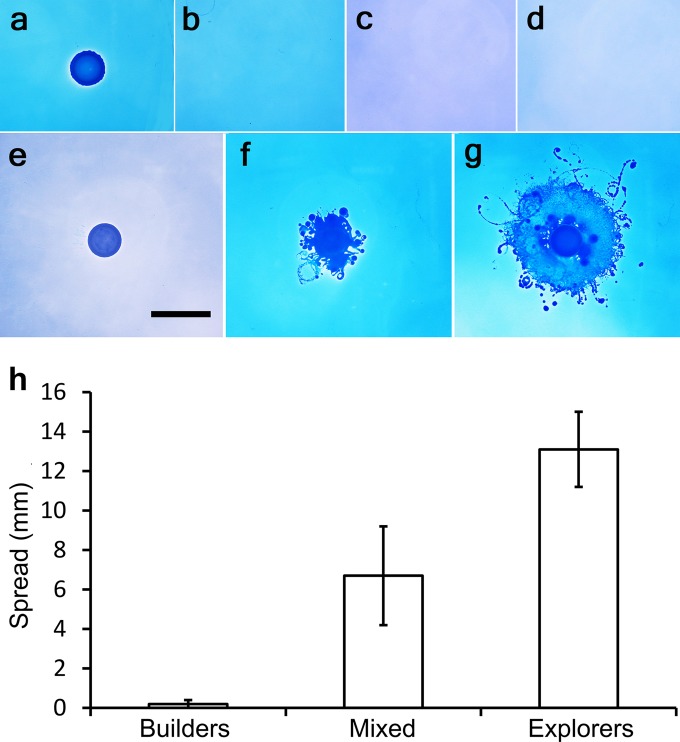
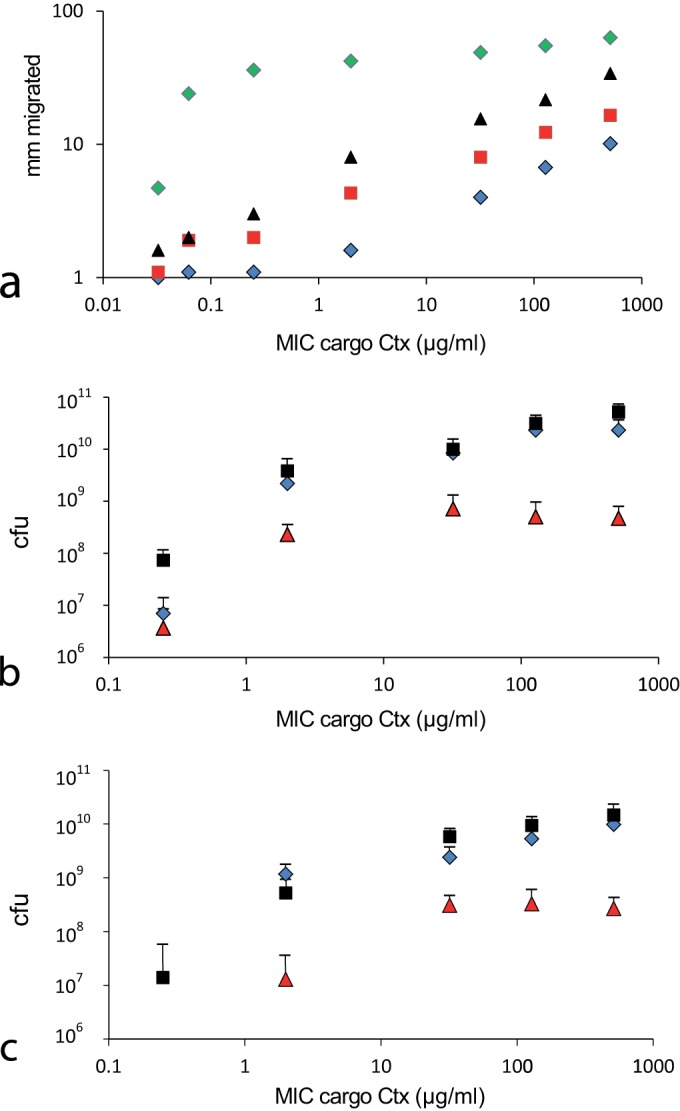
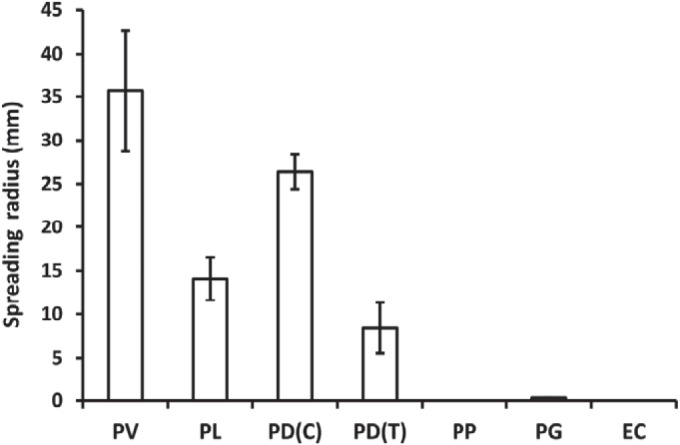
Swarming bacteria are challenged by the need to invade hostile environments. Swarms of the flagellated bacterium Paenibacillus vortex can collectively transport other microorganisms. Here we show that P. vortex can invade toxic environments by carrying antibiotic-degrading bacteria; this transport is mediated by a specialized, phenotypic subpopulation utilizing a process not dependent on cargo motility. Swarms of beta-lactam antibiotic (BLA)-sensitive P. vortex used beta-lactamase-producing, resistant, cargo bacteria to detoxify BLAs in their path. In the presence of BLAs, both transporter and cargo bacteria gained from this temporary cooperation; there was a positive correlation between BLA resistance and dispersal. P. vortex transported only the most beneficial antibiotic-resistant cargo (including environmental and clinical isolates) in a sustained way. P. vortex displayed a bet-hedging strategy that promoted the colonization of nontoxic niches by P. vortex alone; when detoxifying cargo bacteria were not needed, they were lost. This work has relevance for the dispersal of antibiotic-resistant microorganisms and for strategies for asymmetric cooperation with agricultural and medical implications.
Importance: Antibiotic resistance is a major health threat. We show a novel mechanism for the local spread of antibiotic resistance. This involves interactions between different bacteria: one species provides an enzyme that detoxifies the antibiotic (a sessile cargo bacterium carrying a resistance gene), while the other (Paenibacillus vortex) moves itself and transports the cargo. P. vortex used a bet-hedging strategy, colonizing new environments alone when the cargo added no benefit, but cooperating when the cargo was needed. This work is of interest in an evolutionary context and sheds light on fundamental questions, such as how environmental antibiotic resistance may lead to clinical resistance and also microbial social organization, as well as the costs, benefits, and risks of dispersal in the environment.
Copyright © 2015 Finkelshtein et al.
Figures
References
-
- Mobley HL, Belas R, Lockatell V, Chippendale G, Trifillis AL, Johnson DE, Warren JW. 1996. Construction of a flagellum-negative mutant of Proteus mirabilis: effect on internalization by human renal epithelial cells and virulence in a mouse model of ascending urinary tract infection. Infect Immun 64:5332–5340. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
