

Genomic and experimental evidence for multiple metabolic functions in the RidA/YjgF/YER057c/UK114 (Rid) protein family
- PMID: 25975565
- PMCID: PMC4433059
- DOI: 10.1186/s12864-015-1584-3
Genomic and experimental evidence for multiple metabolic functions in the RidA/YjgF/YER057c/UK114 (Rid) protein family
Abstract
Background: It is now recognized that enzymatic or chemical side-reactions can convert normal metabolites to useless or toxic ones and that a suite of enzymes exists to mitigate such metabolite damage. Examples are the reactive imine/enamine intermediates produced by threonine dehydratase, which damage the pyridoxal 5'-phosphate cofactor of various enzymes causing inactivation. This damage is pre-empted by RidA proteins, which hydrolyze the imines before they do harm. RidA proteins belong to the YjgF/YER057c/UK114 family (here renamed the Rid family). Most other members of this diverse and ubiquitous family lack defined functions.
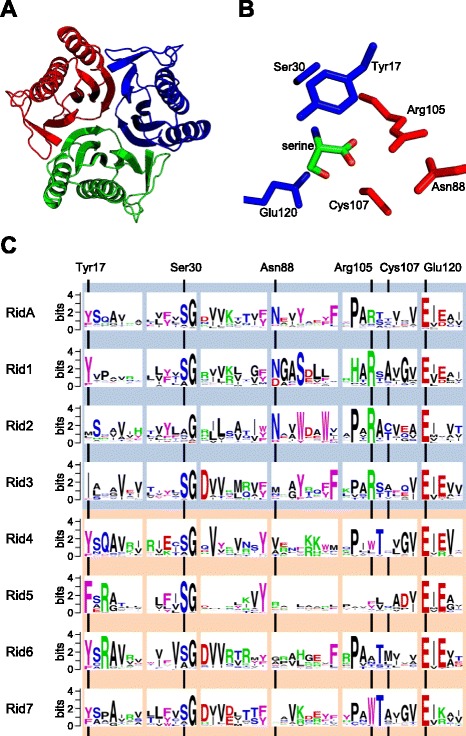
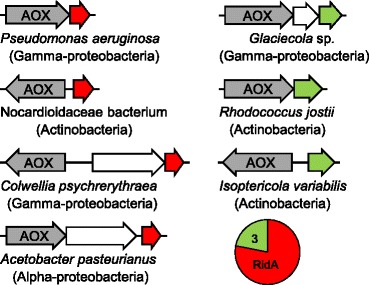
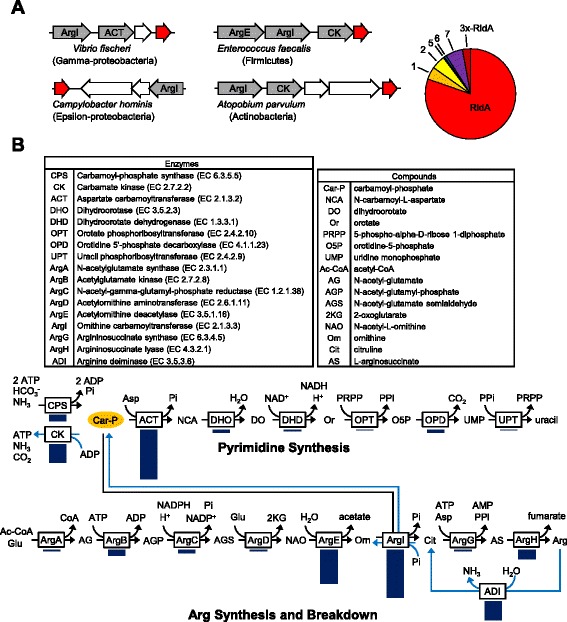
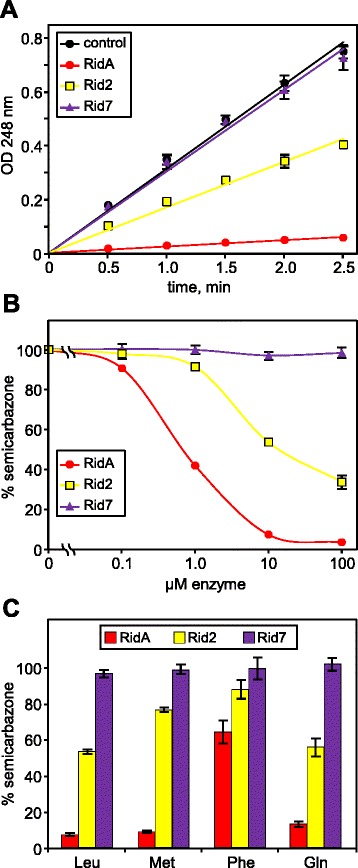
Results: Phylogenetic analysis divided the Rid family into a widely distributed, apparently archetypal RidA subfamily and seven other subfamilies (Rid1 to Rid7) that are largely confined to bacteria and often co-occur in the same organism with RidA and each other. The Rid1 to Rid3 subfamilies, but not the Rid4 to Rid7 subfamilies, have a conserved arginine residue that, in RidA proteins, is essential for imine-hydrolyzing activity. Analysis of the chromosomal context of bacterial RidA genes revealed clustering with genes for threonine dehydratase and other pyridoxal 5'-phosphate-dependent enzymes, which fits with the known RidA imine hydrolase activity. Clustering was also evident between Rid family genes and genes specifying FAD-dependent amine oxidases or enzymes of carbamoyl phosphate metabolism. Biochemical assays showed that Salmonella enterica RidA and Rid2, but not Rid7, can hydrolyze imines generated by amino acid oxidase. Genetic tests indicated that carbamoyl phosphate overproduction is toxic to S. enterica cells lacking RidA, and metabolomic profiling of Rid knockout strains showed ten-fold accumulation of the carbamoyl phosphate-related metabolite dihydroorotate.
Conclusions: Like the archetypal RidA subfamily, the Rid2, and probably the Rid1 and Rid3 subfamilies, have imine-hydrolyzing activity and can pre-empt damage from imines formed by amine oxidases as well as by pyridoxal 5'-phosphate enzymes. The RidA subfamily has an additional damage pre-emption role in carbamoyl phosphate metabolism that has yet to be biochemically defined. Finally, the Rid4 to Rid7 subfamilies appear not to hydrolyze imines and thus remain mysterious.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
